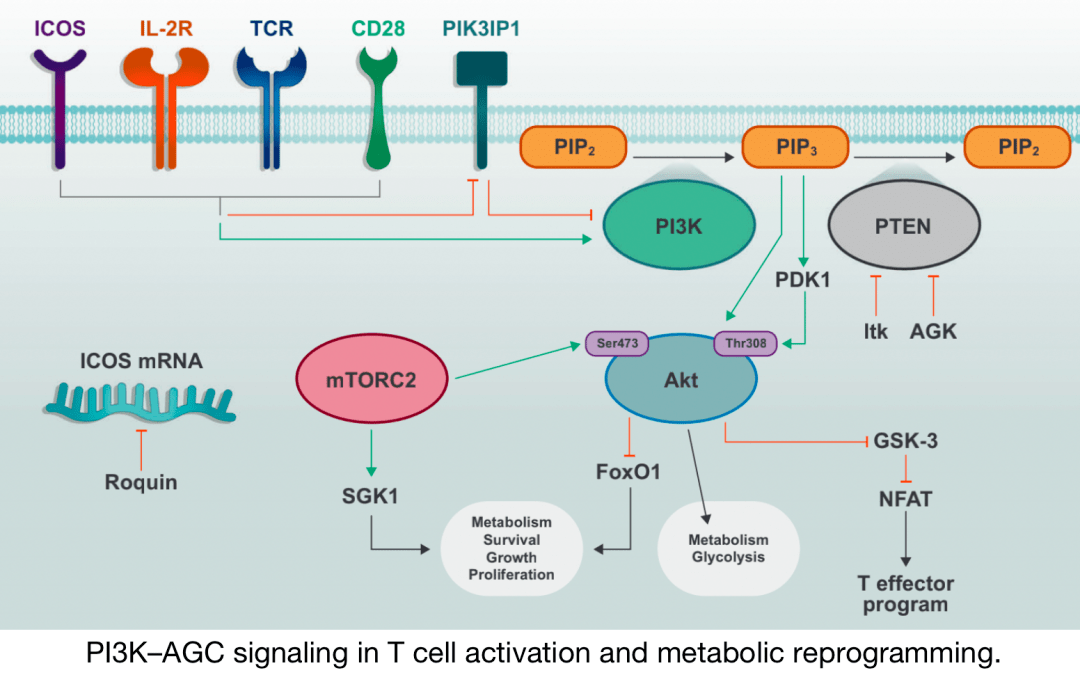
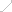��1. PI3K-AGC signaling
��֬��Ӱ���������ߴ�л;���Ĺؼ��ڶ���ʹ�������ת�������ܵ��߶ȵ��ڡ���֬ת����һ�����Ľ�����PI3K������ ��֬������-��4,5��-��������PIP2��ת��Ϊ ��֬������-��3,4,5��-��������PIP3����PIP3�IJ������� ��Ĥļ���� ����PH��pleckstrin homology���ṹ��ĵ����ʹ��ܵ�����
��ˣ�PI3K���Կ���ͨ������������PH�ṹ���ЧӦ������ļ���ڽ�������������ϸ���ź�������PI3K�����յ��������ϸ�����ܵĶ����ź�ͨ·������Akt������øB���� ��֬�����������Ե���ø1��PDK1���Լ� mTOR������1��
PI3K�CTϸ����ʹ�л�ر���е�AGC�ź�
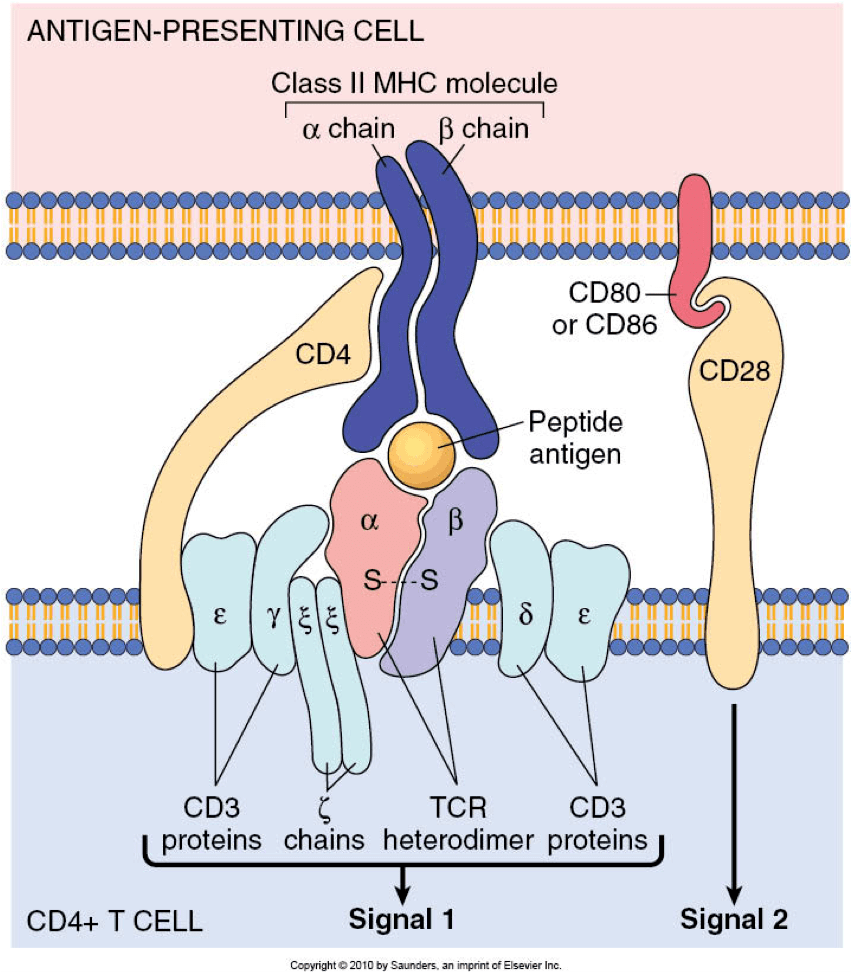
TCR��CD28��IL-2R�ļ����յ�PI3K�����ữ�ͼ������Լ� PI3K���Ʒ��ӵ�ʧ����PIP2ͨ�� PI3K�Ļ���ת��Ϊ PIP3��PIP3�ٽ�����PDK1��Akt���ڵ������źŷ��ӵ� ��Ĥļ���ͼ����� mTORC2��һ�� ����Akt���ٽ���л������ Tϸ��ЧӦ��������
��1.1PI3K
��Protein kinase C��perfectly balanced��
I��PI3K��һ�� ������嵰��������һ������p110�ǵ�λ ��p110����p110�¡�p110�Ļ�p110�ã�����������ǵ�λ������p50��, p55��, p55��, p85��, or p85�£��е�һ����ɡ��� û�д̼�������£� ���ǵ�λ�ļ�ø�ṹ���� �����ǵ�λ������PI3K�����Ũ����ϸ������״̬������أ���Tϸ���У� ϸ������״̬�� TCR��ϡ�CD28����鵼�Ĺ��̼���ϸ�������ź�����IL-2R���Σ��������� ���μ�����PI3K�� Src-homology-2��SH2���ṹ��� �������������νӵ����ϵ� ���������ữYXXM���������µ����ǵ�λ������������ �ͷźʹ��ǵ�λ��λ�������������Ĥ��
PI3K�ź�������ø���Ը�������������ԣ�PIP3�ֱ�ͨ�� ����ø�� ��������ͬϵ����PTEN���Լ� ��SH2�ṹ���� ����5��-����ø��SHIP��ת��Ϊ PIP2�� PI-��3,4��-P2��SHIP�� �ٽ�Th1ϸ����Ӧ������������Ҫ�����ã��� PTENȱ���ᵼ��Tϸ�����Ȼ���������ڴ��Ŵ̼��¡�PTEN��ǿTCR�̼���CD28���̼��±� ����������ԭ���� Tec���弤øIL-2�յ��� Tϸ����ø��Itk����������CD8 Tϸ�������ڼ�Ҳ�ܵ� AGK�鵼�����ữ�ĸ�������
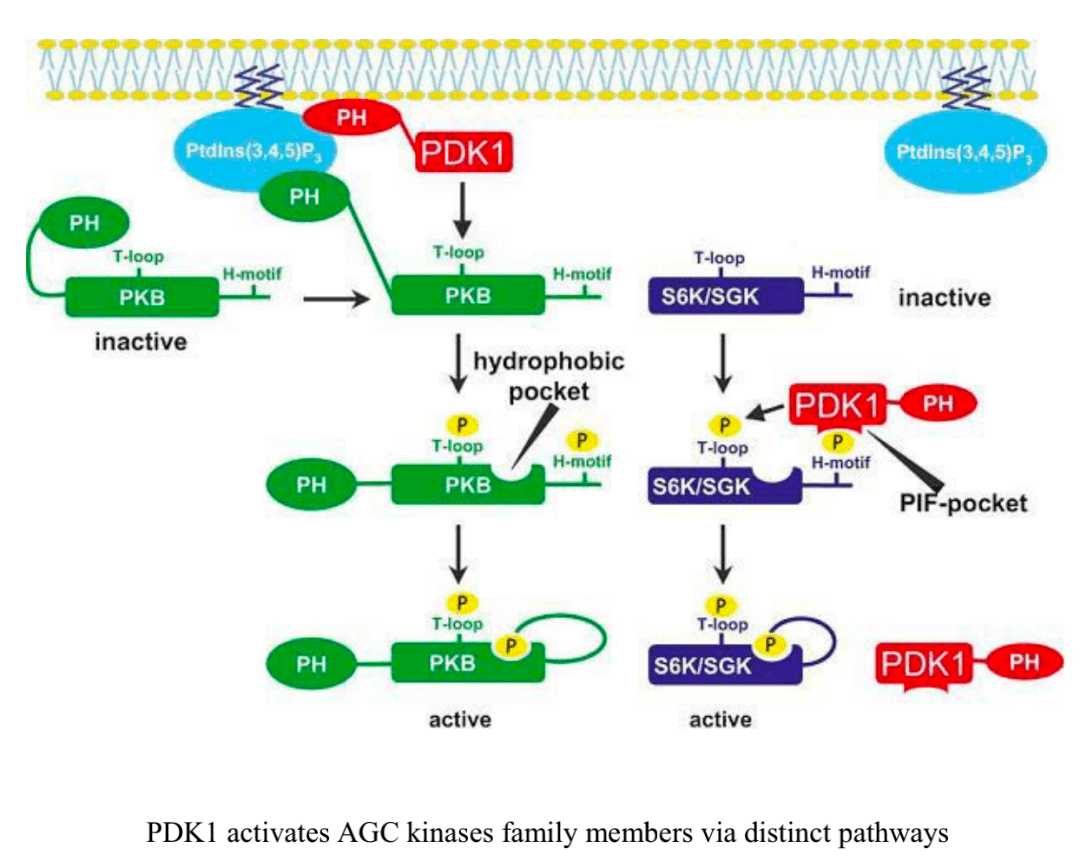
��1.2 AGC��ø
��������������PH�ṹ���ĵ���������AGC��ø�ij�Ա������ PDK1��Akt��������S6��ø��RSK��Ҳ��Ϊp90���� Ѫ��/��Ƥ�ʼ��ص��ڼ�ø1��SGK1����PDK1��һ��˿����/�հ��ἤø���ڼ�������AGC��ø������Akt��SGK1��������Ҫ���á�TCR��CD28���� �ٽ�PDK1����Ĥ��ļ�����յ������ữ�� Thr513�� PDK1�ĵ���øC-����PKC-�ȣ��������������������Ǽ�ø�ṹ��л���Ser241�����Զ����ữ�����������Tϸ���������Ҫ���ڻ��CD8 Tϸ���У�PDK1�����л�ر����ά��IL-2�̼����ε� ��������ȡ���ǽͽ����ù�����Ҫ mTOR�CHIF-1����ȱ���յ�����-1��������������ҪPI3K��Akt���ԡ�PDK1���Ե��ʵ����ڶ�����ǿ�ʵ���Tϸ����Լ�������֢��Tregϸ������������Ҫ�� ��Ser473����mTORC2���ữ�� Akt�ﵽ���� ����Akt��ϵ�����ΪPDK1�ġ�PIF�ڴ����ĵ���dock����ٽ���Thr308�� Akt�����ữ�� mTORC2�CAkt�ź������ Э��Tϸ������ֻ���������
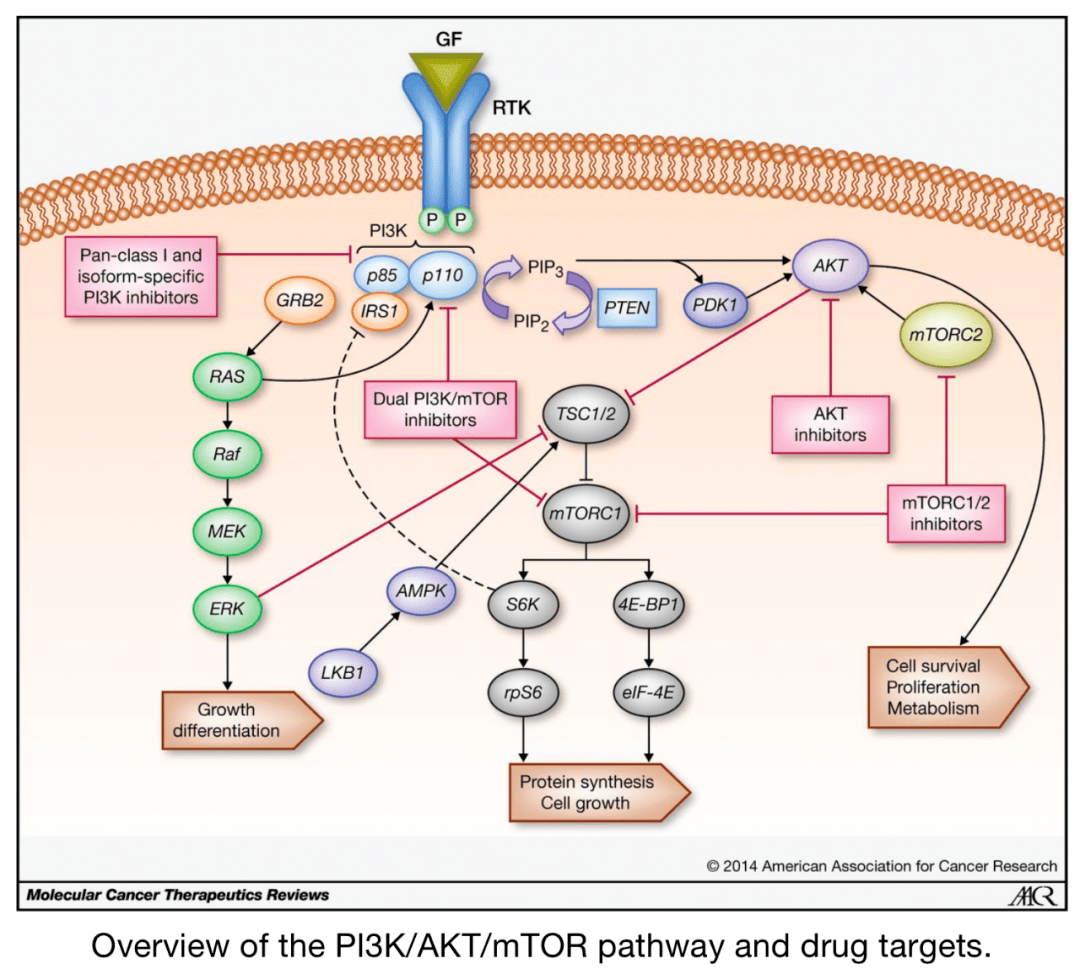
��The Clinical Implications of the Survival Pathway in Prostate Cancer��
Akt��Tϸ���е�һ����Ҫ������ ͨ�����ữ���ڲ�ͷ��O��FoxO��ת¼�����Ļ��ԣ�������� ���ų���ϸ����֮�Ⲣ��ֹ�л���ת¼�� FoxO�����ھ�ֹ��ϸ��Ⱥ�и���ת¼���ԣ�����ͨ��KLF2�ٽ���̬ϸ����������IL7R�ı����Լ�ϸ�������������CD62L��CCR7��S1PR1���ı�������֤�ݱ���FoxO1���Ե��ʵ��µ���Tϸ���ڻ����ȶ�������Ҫ���ӻ����Ͻ����Tϸ���е�FoxO1�µ�����Э��ϸ����������ֳ�dz���Ҫ����Ϊ������������mTORC1�źźͺϳɴ�л����ˣ�Akt�CFoxO1�ᶯ̬����Tϸ����Ӧ��
��һ�ֲ���Akt�źŴ�����˿����/�հ��ἤø����ԭ�ϳ�ø��ø3��GSK-3�������� ��Ϣϸ���о�������Ի��ԣ���ͨ��Akt�����ữ��ʧ�����GSK-3��T��Bϸ����ͨ�����ƻTϸ���ĺ����� ��NFAT�����ԣ��ٽ�������ƻ�� AGC��ø����RSK���� �������Tϸ����л�滮�е�ȷ������һֱ���ܹ�ע��Ҳͬ����Ŀǰ�о����ص㷽��
��2. mTOR signaling
mTOR��һ��˿����/�հ��ᵰ��ø�������� �����źŸ�����mTORC1��mTORC2���� mTORC1��mTOR��ø���壬mTORC1�����䵰������ص���SEC13����8��MLST8���IJ��鶯���������ף��Լ�������Akt����-1��PRAS1�������DEP�ṹ������õ�����DEPTOR���������ǵ�λ��mTORC1�źŶ� �����е�Tϸ�����������ܵ��ڻ����ȶ��ͷֻ�ΪЧӦCD4 Th1��Th2��Th17ϸ���Լ�ϸ������CD8 Tϸ��������Ҫ�� ���֮�£� Th1��Th2ϸ���ֻ�����ҪmTORC2���ԣ�ͬʱ����Ҳ�� ����Tfh��Tregϸ����Ǩ����
mTOR�ź�ͨ����������Tregϸ���ļ����ϵ�ȶ��Ժ����ƹ������� �Կ���ͳTϸ����Ӧ�з��Ӷ������������mTOR�źſ����ܰ���֯�� ���ڼ���CD8 Tϸ���ķֻ���ȴ�� �ٽ����ڷ��ܰ���֯�еķ�������ˣ�mTOR�ź���Tϸ����Ӧ�� �����������
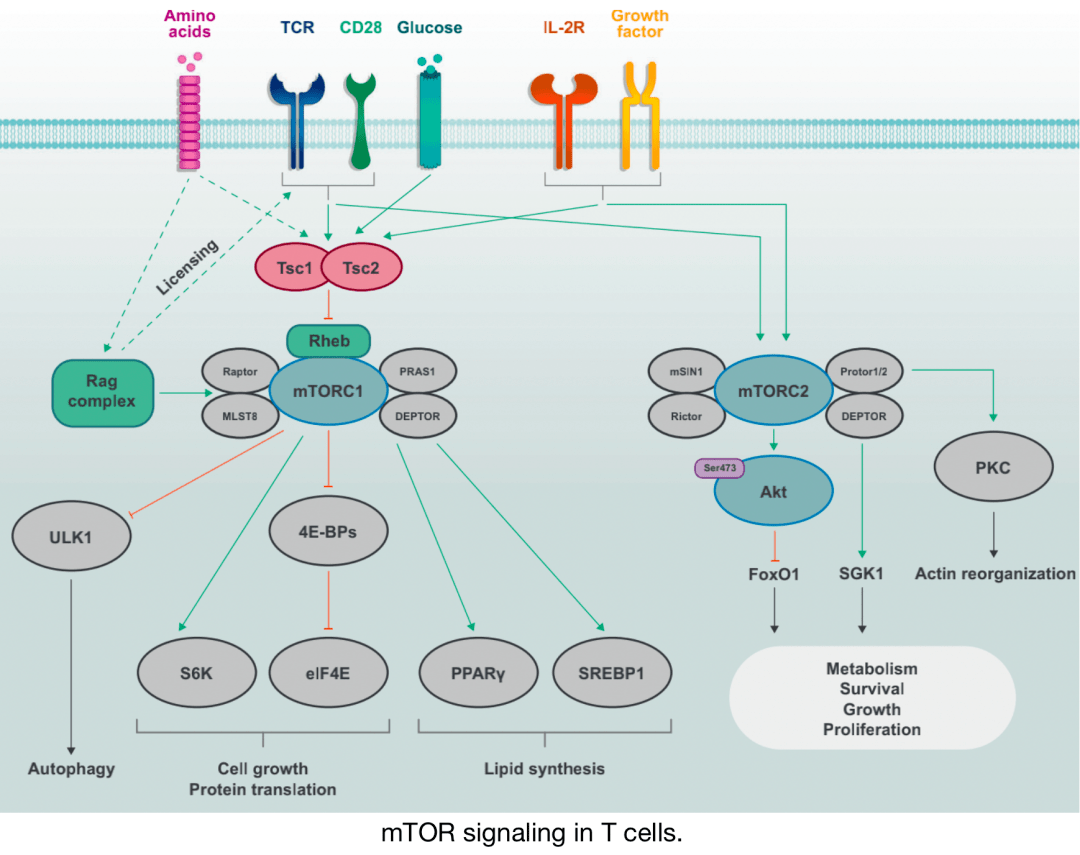
Tϸ���е�mTOR�ź�
��ɢ��mTOR������mTORC1����mTOR��Raptor��PRAS1��DEPTOR��MLST8��ɣ���mTORC2����mTOR��Rictor��Protor1/2��mSIN1��DEPTOR��ɣ�������������TCR��CD28��IL-2R�����������Ӽ��mTORC1�ļ���Ҳ�������Ӫ����������������ͨ��Rag������ٽ�mTORC1���Rag������Ҳ������TCR��CD28���̼��ź��յ�mTORC1����ġ����ɡ����á�Tsc�������Ļ����ܵ����ߺ����������źŵ����ƣ�ͨ������СG����Rheb�ļ�����ߴٽ�mTORC1�ļ��mTORC1ͨ��S6K��eIF4E�յ�ϸ�������͵����ʷ��룬��ͨ��PPAR�ú�SREBP1�յ�֬�ʺϳɡ���Ӫ������������£�mTORC1ͨ��ULK1�������ɡ�mTORC2��Ҫ��ϸ����������ֳ�ʹ��Ļ������������Դ�л�ر�������Źؼ����á�
��3. LKB1-AMPK signaling
LKB1�CAMPK�ź�ͨ·�ڵ���ϸ����л����ֳ��������Ӧ��Ӫ������������ĸı䷽�����ź���������LKB1�CAMPK�źſ� �ٽ�����ATP�ķֽ��л;������ʹTϸ����Ӧ�� ����Ӧ��ʱ���д�л��������ͨ�����ڴ�л�ر�̣�LKB1��AMPK������ Tϸ���ķֻ������������²��֣����ǽ�����LKB1��AMPK��������ε��ڵģ���Դ�л��Ӱ���Լ���Tϸ���鵼�������е����á�
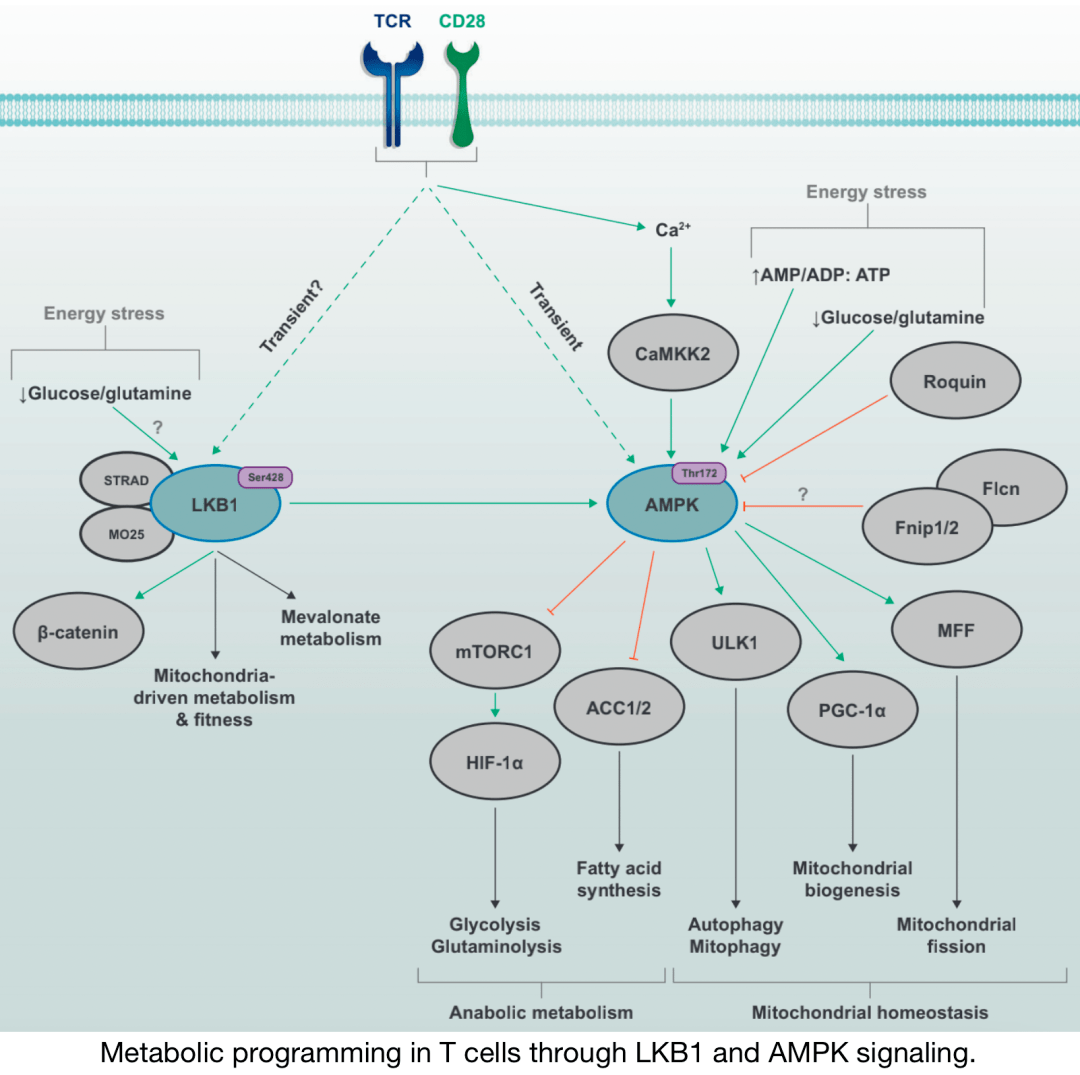
ͨ��LKB1��AMPK�ź���Tϸ���н��д�л���
����Ӧ��;����øLKB1��AMPK��TCR��CD28���̼��źż�� AMPK���Բ����� Ca2+CAMMK2;���鵼���� ȱ�������ǻ�Ȱ�������AMP/ADP/ATP��ֵʧ����Ҳ�� �ٽ�LKB1�CAMPK�źŴ��������� Ӫ����Ӧ��������Fnip�CFlcn�������Roquin������ ����AMPK������ LKB1�ļ������������л����Ӧ�ȵı仯�Լ��ض������� ����������л�������йء�ͨ�����ڶ�����ΰе�Ļ��ԣ�AMPK�źſ� ��ֹ�ǽͽ⡢�Ȱ��������֬����ϳɵĴ�л����ͬʱ�ٽ��ֽ��л����������˿�������ɺ����ɡ�AMPK��ͨ�� �ٽ����������������������������������嶯��ѧ���Ӷ�֧����������ԡ�
��3.1 LKB1��AMPK�ĵ���
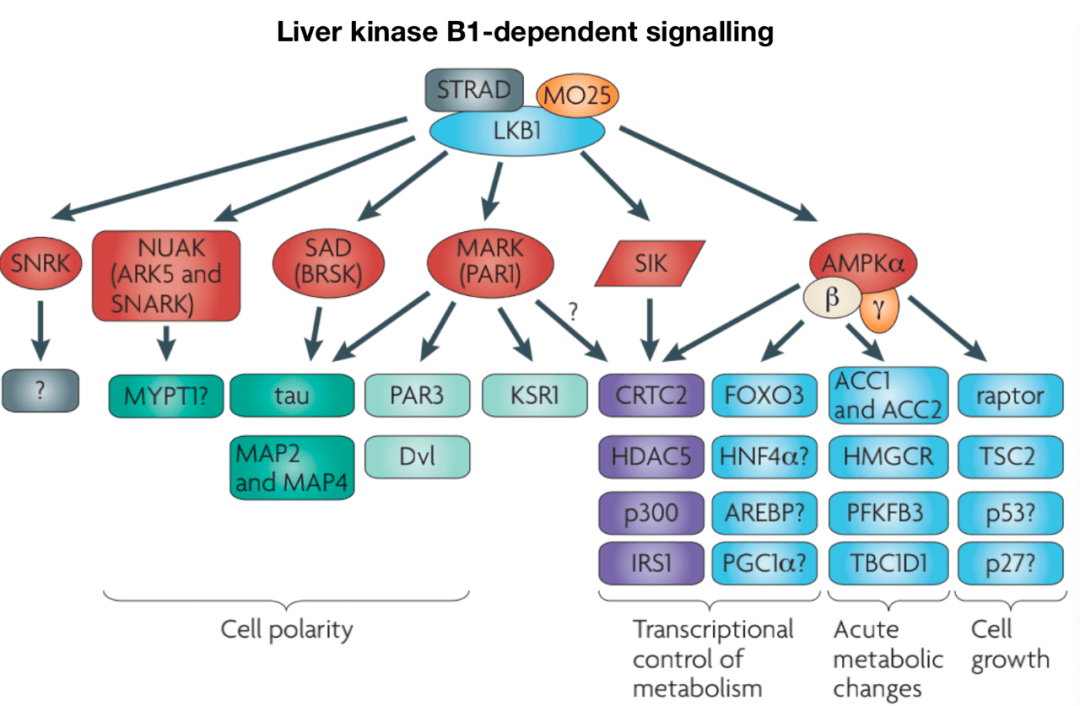
��The LKB1-AMPK pathway��metabolism and growth control in tumour suppression��
LKB1��һ��˿����/�հ��ἤø�������������ƹ��ܣ��������ϸ����л����ֳ��LKB1 ���������ΰе���������������ȷ�İе� AMPK��AMPK��ؼ�ø����BRSK��NUAK��MARK��LKB1�����ε�����ϸ����λ�ͷ�������ν鵼��LKB1 ��STE20�����������STRAD����MO25�γɸ��������ֱ�ٽ�LKB1�� ϸ���ʶ�λ�ͼ�ø���������⣬ Akt����ͨ���ٽ�LKB1������������LKB1��
AMPK��һ�ֱ��ص�˿����/�հ��ἤø���ɦ����ºͦ��ǻ���ɡ�AMP��ADP��CBS3�ظ����еĽ������������ø��AMPK�����ǻ���Thr172�ķ��ʣ����ǻ��Ǵٽ�AMPK��ø���ԵĹؼ����ữλ�㡣���գ� ��л����ĸ��ӵ�������AMPK�����ڲ�ͬ����Ӧ�������µġ��ּ�����Ӧ�������������μ�ø�鵼AMPK Thr172���ữ�� LKB1���ƸƵ��ؼ�ø��ø2��CaMKK2���� TGF-�¼��ø1��TAK1��������LKB1��CaMKK2��������Ϊ�������Tϸ���У�TCR��Ca2�ź��Լ�CD28���̼�Эͬ���ã�ͨ��CaMKK2���١�˲ʱ����AMPK��AMPK����Ҳͨ�� ������-��������������е��ڡ� Fnip1��Fnip2��AMPK�γɸ�������Fnip1�Ĺ���ȱʧͻ�������Bϸ����AMPK���ԣ��Ӷ�ȷ��Fnip1��AMPK�ĸ��������ӡ�Fnip��ϵ���FlcnҲ��AMPK����ã�������ΪAMPK�źŵĸ��������ӣ���֤�ݱ���Fnip1��Flcn��AMPK������ڡ�
�� 3.2 LKBI-AMPK�鵼�ķֽ��л
AMPK��������Ӫ������յ�һ�£�ͨ�����ֽ��л�����йأ�������ATP��AMPKͨ��ULK1�����ữ�������ɣ�ULK1�ɴٽ����ɽ鵼������������̬���ٽ�Tϸ�������⣬AMPK������mTOR���ڣ�������������ʱ��AMPK��ͨ��Ser15��p53�����ữ�鵼ϸ���������͡���ˣ�LKB1�CAMPK�ź�ͨ����������Ӧ���ڼ�ĺϳɴ�л��ϸ�������ٽ�ϸ����������⣬ͨ����������;�������Ĵ�л���ͪ-5-�����LKB1�CAMPK;�����ƻ��ٽ���֬�����ɡ��෴��LKB1Ҳ���Դٽ����������л����AMPK�������Է�ʽ֧��Tregϸ������̬����ˣ�LKB1�CAMPK�ź���Ҫ����������֬�ʺϳ�;������AMPK������LKB1�źſ�����ѡ��������´ٽ�����֬��������
������ϸ�����о�������LKB1��AMPK�źŵ��ƻ��ٽ������ǽͽ���������ͨ��HIF-1������������ǽͽ�ø��ת¼��HIF-1����LKB1�CAMPK�����Ե���Ҳ���ܲ���������mTORC1�����ơ�LKB1�CAMPK�źſ���ͨ��HIF-1����ACC1�鵼���ǽͽ��������������л�仯���Э��Th17��Tregϸ��ϵ�ķֻ������⣬������о�������LKB1�ٽ��ȶ���Foxp3����Լ�Th2��Tregϸ���ķ�����������AMPK��mTORC1�CHIF-1���źţ��������ڦ�-���������źš�LKB1�ź���TCR�鵼��Tregϸ������������幦�ܺ������������Դ�л����������ģ�����FAO�����ʺ���ऴ�л����Щ����ǿ��LKB1��AMPKЭ����л�ر���Ե���Tϸ���ֻ���Tregϸ��������
��4. ���ߴ�л�ź�����
������Ҫ�ź�ͨ·�ڹ����ϲ����ų⣬�෴���������֯�����������ͬ�ﵽ�ʵ��� ��л������������ϸ�����ܵ��ض�����������ˣ�������Щ �����������ڹ�ͬ�Ľڵ��ϣ����� �������Ӻ�����ϸ��������
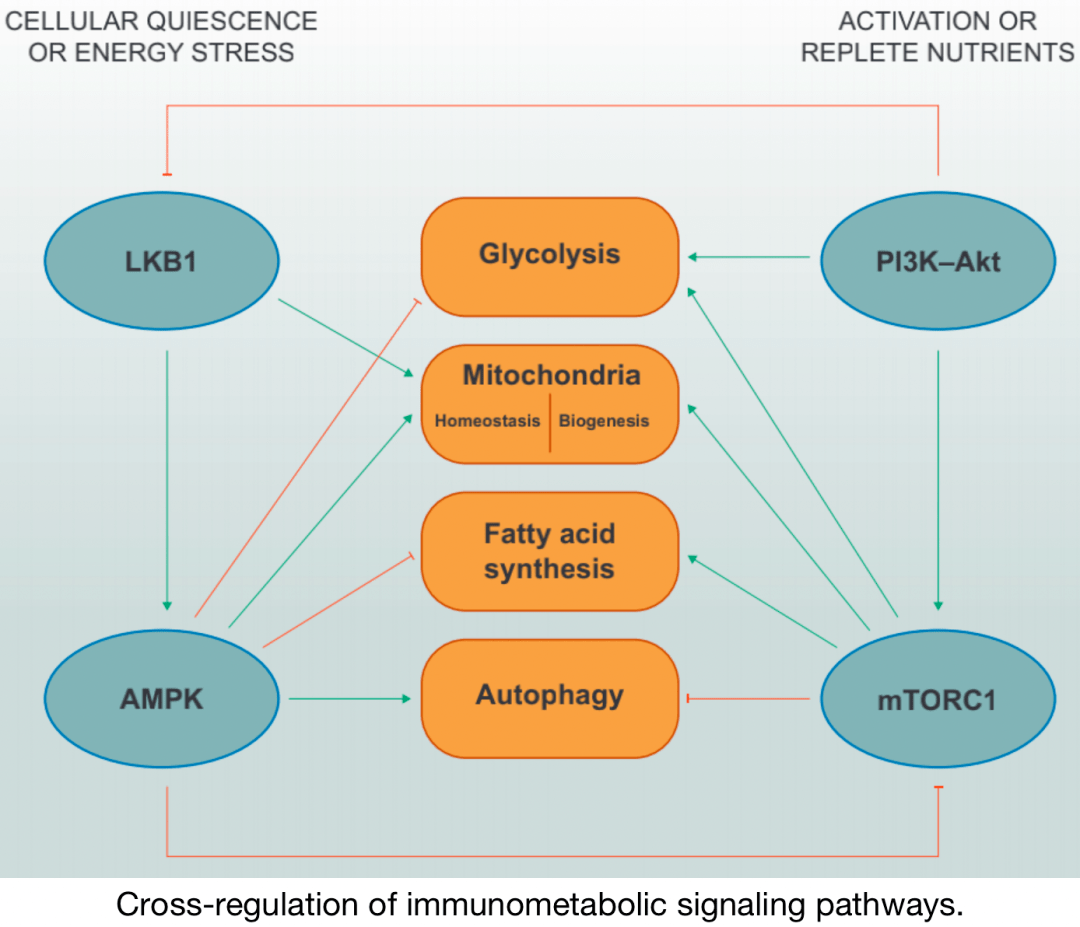
���ߴ�лͨ·�Ľ������
�� Ӫ��ȱ���������£� LKB1�CAMPK�ź� ���ƺϳ���л��س������ǽͽ��֬����ϳɣ�ͬʱ�ٽ�����������̬�����ɡ�AMPKͨ����רһ��ת��������Raptor�����ữֱ������mTORC1���� �����/��Ӫ�������������£� PI3K�CAkt��mTORC1�ź� �ٽ��ǽͽ⡢�������������֬���� �ϳ���ͬʱ �����������ݱ����� Akt�������ữLKB1�� �����书�ܶ�λ��
һ����˵�� PI3K�CAGC��ø��mTOR���Ե����ߣ��Լ�LKB1�CAMPK���ԵĽ������� ϸ����������ֳ��ЧӦ��������أ������л����ı仯��ء� ���θ�������ź��������������δ�л����ĵ����й�����������ϸ����л������ź�ͨ·Ҳ�ܵ���л���Ӫ�����ʵĽ���Ӱ����������˫���л�źš�Ϊ��ø�������źźͻ���ת¼�ṩ����һ����أ��� ���Ʋ�ͬ�����µ�����ϸ��������
�����ź����������ߴ�л�е�������һ�������˷ܵ��о��������ƺ����ཡ�����й㷺�����塣����ƪ�����У�����ǿ����PI3K�CAGC��mTOR��LKB1�CAMPK�ź���Tϸ�����ܺ������еĹؼ���������������������Щ�źŷ�������ε��ڵ��Լ�������������±��Tϸ����л����ȡ���˺ܴ��չ��ȷ����� ֪�������������ź�����TCR��ϸ���������壩����ڴ�л�д��ݲ�ͬ�Ĺ��ܳ���������Ϊ ���ƹ�����Tϸ���������翹����CAR-Tϸ�����ƣ��ṩ�µ����ưе���
�ο����ף�
Saravia, J., Raynor, J.L., Chapman, N.M. et al. Signaling networks in immunometabolism. Cell Res 30, 328�C342��2020��. https://doi.org/10.1038/s41422-020-0301-1