疼痛是肿瘤最常见的并发症之一。在接受治疗的肿瘤患者中,超过1/3的患者会遭受疼痛折磨[1]。
有研究发现,疼痛会抑制免疫反应,促进肿瘤的生长[2]。不过,疼痛、肿瘤及免疫三者间的联系非常复杂[3],还有很多谜团有待科学家解开。
近日,蒙特利尔大学Sebastien Talbot带领的团队,在《自然》杂志上发表了伤害感受神经元促进肿瘤生长的相关研究[4],让我们对神经和抗肿瘤免疫有了新的认知。
他们发现,黑色素瘤细胞会激活伤害感受神经元,诱导伤害感受神经元释放神经肽CGRP,导致CD8+T细胞进入耗竭状态,而阻断这一通路可以破除肿瘤的免疫逃逸。
 论文首页截图
论文首页截图血管生成对肿瘤进展的促进作用已广为人知[5]。近年来,随着神经科学相关技术的发展,人们逐渐认识到神经系统对肿瘤也有重要影响。
例如,交感肾上腺素能神经可激活肿瘤中血管内皮细胞,从而促进肿瘤生成[6,7]。此外,交感神经也可抑制免疫系统,辅助肿瘤细胞逃逸[8]。除了外周自主神经,中枢神经系统中的神经祖细胞可诱导肿瘤内神经生成并驱动肿瘤生长[9]。
伤害感受神经元对肿瘤的神经支配是否影响肿瘤免疫,目前仍有待研究。
Sebastien等将表达GFP的B16F10黑色素瘤(其肿瘤细胞带绿色荧光标记)接种在Nav1.8Cre::tdTomatofl/WT小鼠上(其伤害感受神经元带紫色荧光标记)。
接种后的第21天,他们在黑色素肿瘤组织中发现大量的伤害感受神经元(图1)。同时,他们在细胞实验中也发现黑色素肿瘤细胞能促进神经元轴突生成。
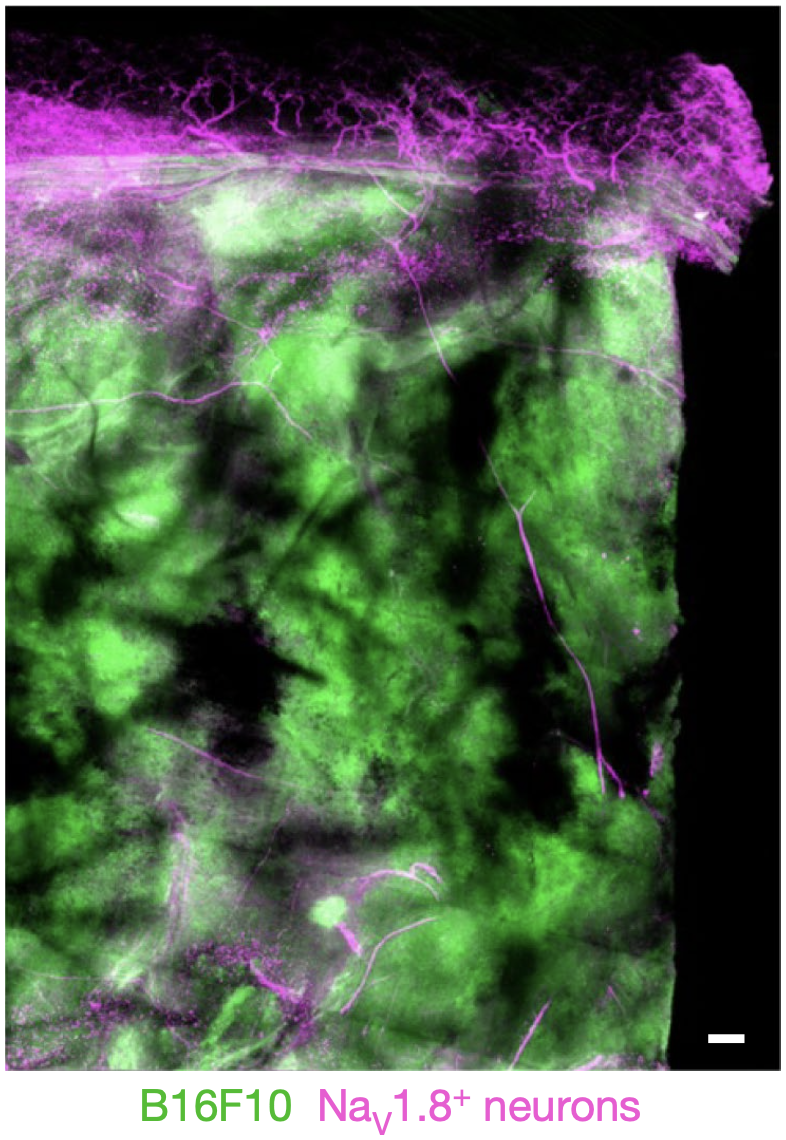 图1. 黑色素瘤中伤害感受神经元的分布
图1. 黑色素瘤中伤害感受神经元的分布为了探索黑色素肿瘤细胞与伤害感受神经元间更深层次的关系,Sebastien等系统地实施了一系列细胞实验。
他们发现,单独培养伤害感受神经元不会释放神经肽,但将其与黑色素肿瘤细胞共培养后,伤害感受神经元被激活(图2A),并释放大量具有免疫调节活性的神经肽,如VIP、SP及降钙素基因相关肽(CGRP),其中CGRP的浓度最高(图2B)。
这个意外的发现促使他们对与黑色素瘤细胞共培养的TRPV1+伤害感受神经元进行了RNA测序。结果显示,伤害感受神经元中编码CGRP的基因Calca和与癌痛相关的基因Trka明显上调(图2C)。上调的Trka可提高神经元对疼痛的敏感度,并促使活化的神经元释放CGRP,这可能是神经元接受癌痛信号后调节免疫系统的潜在方式。
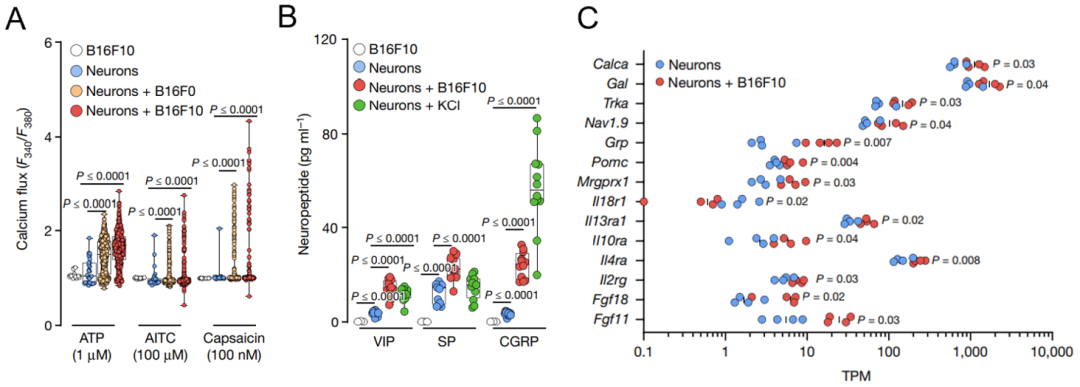 图2. 黑色素肿瘤细胞激活伤害感受神经元
图2. 黑色素肿瘤细胞激活伤害感受神经元为了进一步探究黑色素瘤细胞激活伤害感受神经元及其免疫调节的机制,Sebastien等进行了系列细胞实验,将肿瘤细胞、CD8+T细胞及神经元三类细胞单独培养或共培养。
实验结果显示,与单独培养的肿瘤细胞相比,与CD8+T细胞或神经元共培养可上调黑色素瘤细胞中Slpi的表达(图3A)。特别是在三类细胞共培养48h后,培养基中SLPI的浓度竟比单独培养肿瘤细胞时高200倍(图3B)。
作为一种分泌性白细胞蛋白酶抑制因子,SLPI不仅可以促进视网膜干细胞的轴突再生,还能诱导神经干细胞增殖[10]。然而,SLPI如何作用于伤害感受神经元呢?
为了回答这个问题,Sebastien等利用钙离子显微镜检测SLPI能否直接激活伤害感受神经元。结果显示,SLPI(0.01-10 ng/ml)可激活至少20%的神经元(图3C)。前期结果表明活化的伤害感受神经元会释放CGRP(图2B),据此,他们推测SLPI可能是黑色素肿瘤细胞激活伤害感受神经元的关键媒介。
黑色素瘤细胞分泌的SLPI浓度为5 ng/ml,而正是5 ng/ml的SLPI可诱导神经元释放CGRP,而非0.1或1 ng/ml(图3D)。据此,Sebastien等得出结论,黑色素肿瘤细胞通过分泌SLPI激活伤害感受神经元,并诱导其释放CGRP。
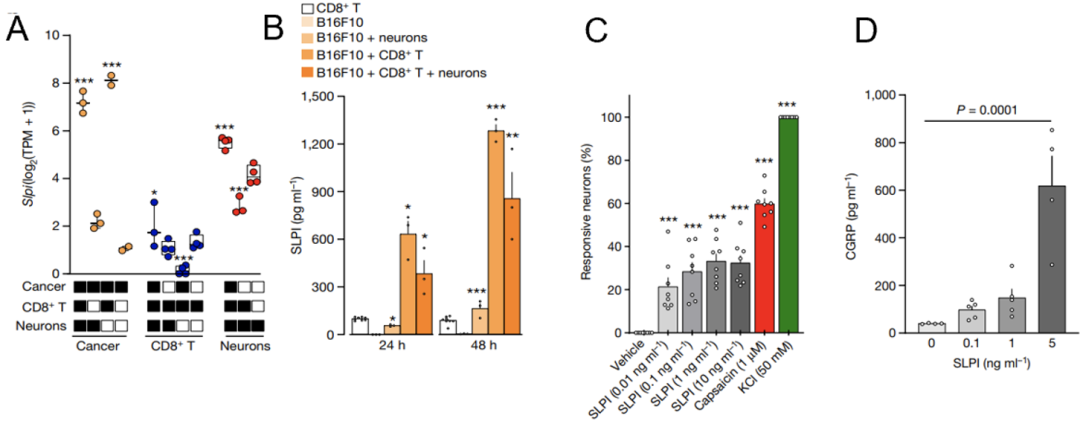 图3. SLPI诱导伤害感受神经元释放CGRP
图3. SLPI诱导伤害感受神经元释放CGRP上述细胞实验证明黑色素瘤细胞能诱导伤害感受神经元释放CGRP,那在肿瘤微环境中这些具有免疫调节活性的CGRP是如何影响免疫细胞呢?
Sebastien等建立了黑素色瘤模型,他们发现荷瘤小鼠对疼痛敏感度与肿瘤浸润的耗竭性CD8+T细胞数量呈正相关(图4A)。将小鼠调控痛觉的基因Trpv1敲除后,肿瘤生长速率下降,且荷瘤小鼠的生存期更长(图4B)。
上述荷瘤小鼠肿瘤组织的流式分析结果显示,Trpv1敲除小鼠的肿瘤中杀伤性IFNγ+CD8+T细胞更多,耗竭性PD1+LAG3+TIM3+CD8+T细胞更少(图4C)。相应地,去除CD8+T细胞后,Trpv1敲除抑制肿瘤生长的作用被逆转了(图4D)。上述结果说明,敲除Trpv1的抗肿瘤作用是依赖于CD8+T细胞。
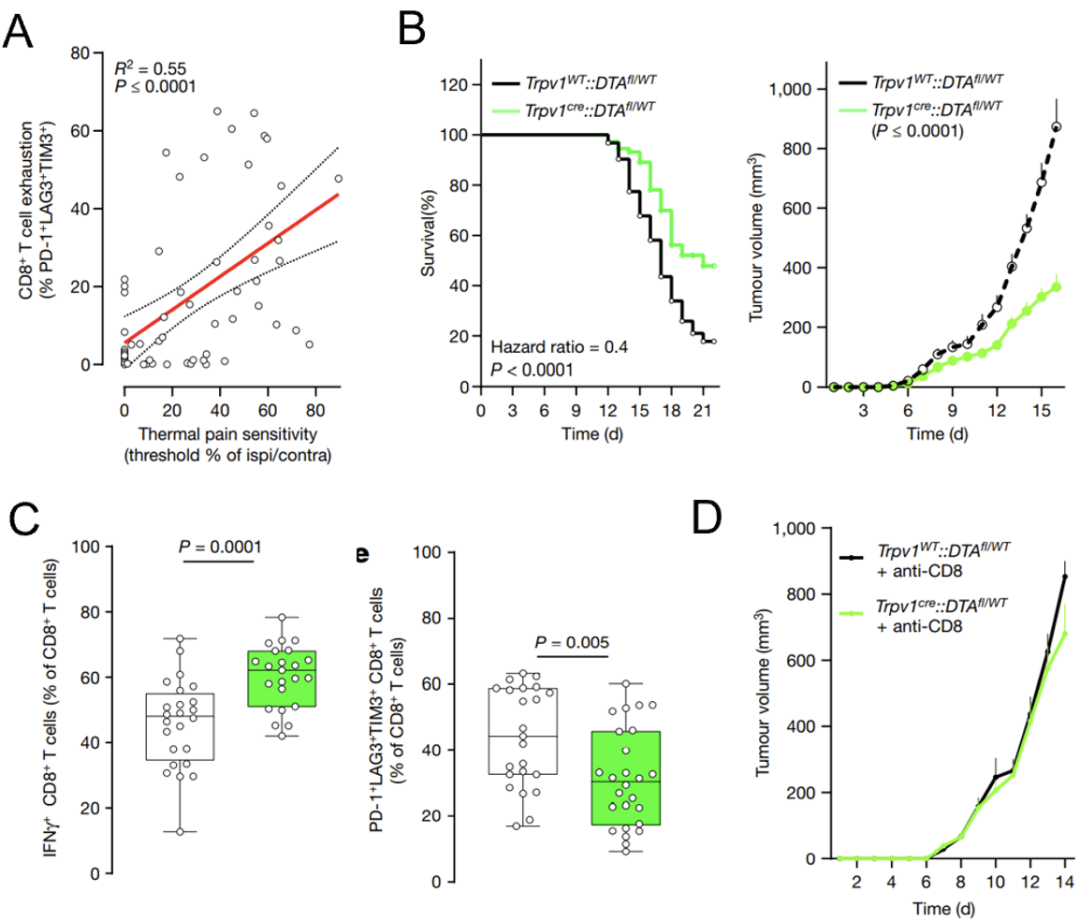 图4. 消融伤害感受神经元可增强抗肿瘤免疫
图4. 消融伤害感受神经元可增强抗肿瘤免疫此前有研究报道,伤害感受神经元释放的神经肽可抑制与CD8+T细胞的抗细菌免疫[11]。Sebastien等也发现CD8+T细胞会表达CGRP的受体RAMP1,那么CGRP-RAMP1轴是否是诱导CD8+T细胞耗竭的关键信号通路?
Sebastien等发现,在野生型小鼠中CGRP会诱导CD8+T细胞进入耗竭状态,降低其对黑色素肿瘤细胞的杀伤力;而在Ramp1敲除小鼠中,CGRP失去了诱导CD8+T细胞耗竭的能力(图5A-B)。
前期研究表明,敲除Trpv1可抑制黑色素肿瘤生长(图4B),而添加CGRP则抵消了Trpv1敲除带来的抗肿瘤效应(图5C)。根据上述数据,他们推测CGRP与RAMP1结合诱导了CD8+T细胞耗竭。
为了确认这一推测,Sebastien等给荷瘤小鼠注射RAMPI的拮抗剂。他们发现这可有效抑制黑色素瘤的生长,同时保持了CD8+T细胞的免疫活性(图5D)。
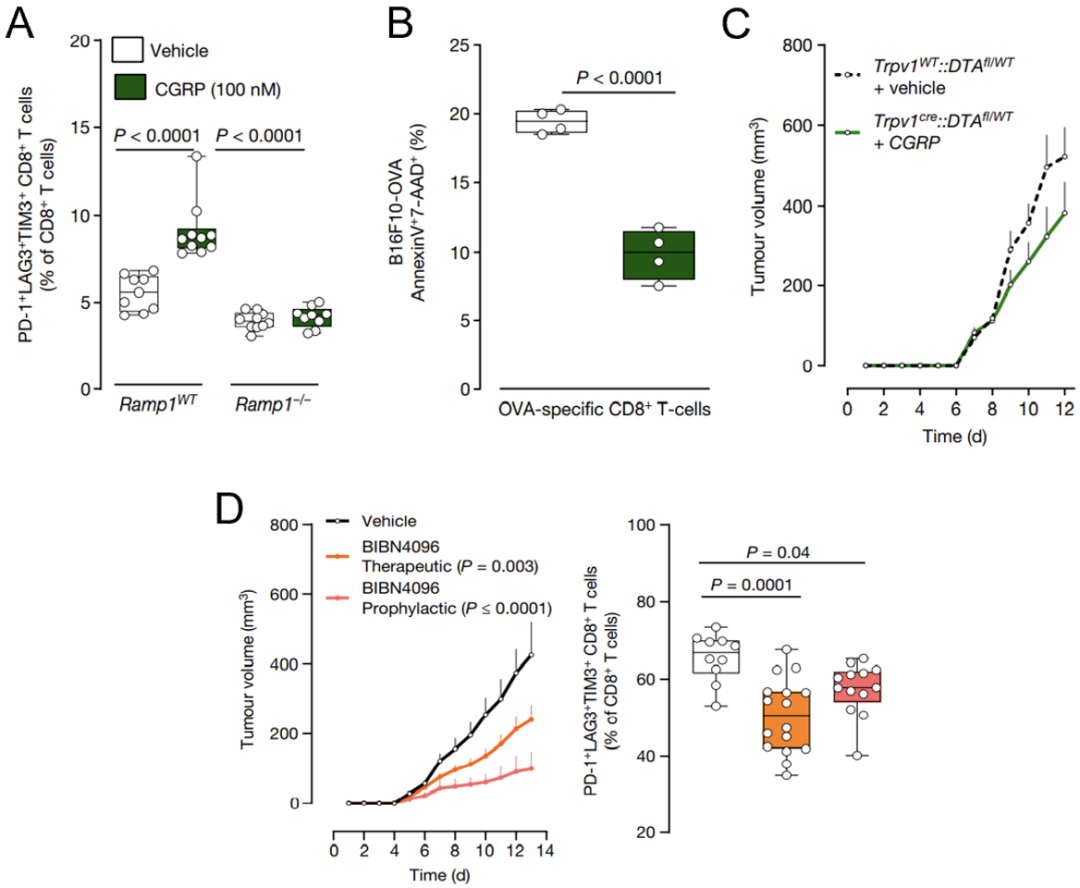 图5. CGRP驱动RAMP1+CD8+T细胞耗竭
图5. CGRP驱动RAMP1+CD8+T细胞耗竭为了进一步说明CGRP-RAMP1通路的重要性,Sebastien等建立了Rag1敲除小鼠的肿瘤模型,然后向其移植野生型或Ramp1敲除小鼠的CD8+T细胞。
他们发现,移植Ramp1敲除小鼠的CD8+T细胞能更好地抑制黑色素瘤生长(图6A)。而且,其免疫活性也较移植野生型小鼠的CD8+T细胞的更高(图6B)。也就是说,敲除Ramp1可保护CD8+T细胞的免疫功能,使其免受RAMP1诱导进入耗竭状态。
进一步,他们分别对野生型与Ramp1敲除小鼠的肿瘤浸润CD8+T细胞进行了RNA测序。结果显示,Ramp1敲除小鼠的CD8+T细胞中免疫抑制性信号分子明显下调,如Pd1、Tim3、Tox及Eomes。上述结果再次确认了RAMP1可直接诱导肿瘤浸润的CD8+T细胞功能耗竭。
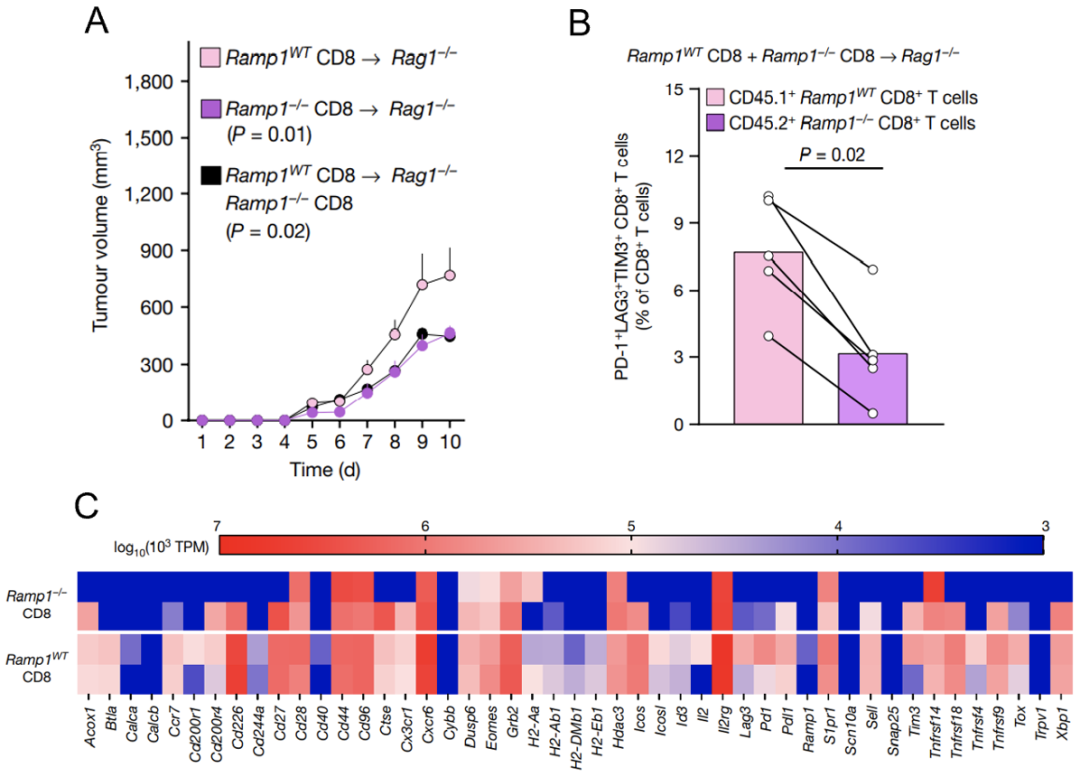 图6.RAMP1诱导CD8+T细胞功能耗竭
图6.RAMP1诱导CD8+T细胞功能耗竭以上结果在黑色素瘤患者中也得到证实。临床数据显示,患者的肿瘤组织中RAMP1表达量越低,他们的生存期就更长(图7A)。
患者肿瘤组织中浸润的CD8+T细胞RNA测序结果显示,高表达RAMP1的CD8+T细胞同时会高表达一些免疫抑制性分子,如PD1,TIM3,LAG3及CTLA4。这些数据说明,RAMP1是肿瘤患者预后不良的标志物。
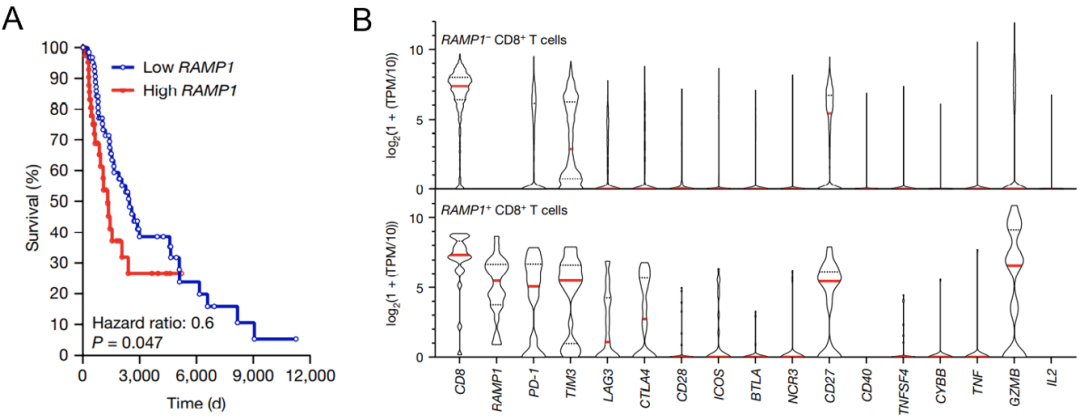 图7. RAMP1是肿瘤患者预后不良的标志物
图7. RAMP1是肿瘤患者预后不良的标志物物实验相结合,Sebastien等证实黑色素肿瘤细胞分泌的SLPI驱动伤害感受神经元释放CGRP,进而通过CGRP-RAMP1信号轴诱导CD8+T细胞耗竭,从而促进黑色素肿瘤生长。
他们发现,活化伤害感受神经元分泌的神经肽是感觉神经促进肿瘤生长的关键纽带,深入揭示了肿瘤细胞、伤害感受神经元以及CD8+T细胞三者间的相互作用。