����2007��ع����¶����������Լ�֮ǰ��������ͬ��Ҳ���������ڲ�ͬ���о���
��ʱ���˱��淢��ƪ���£�����ʱ�侫������̽�����̿��˼��������о���
ͬʱ���پ�����ִ��Ŵ�ѧ���������Ժ˴Ź�������о��������֪��ʼ��2007��ع�֮ǰһ�η��ʱ���ʦ����ѧ���˺֣�������ǧ��������ߣ����м�����ϵ����һ����ʮ�˵Ĵ��ϵ����18λ��ä�ߣ������µ�2024��3��18�ձ�Genetics��־���ܣ�3��28�����Ϸ�����
����ʶ���Ǹ������֪��Ҳ�Ǹ��Ӿ���֪��
��ʶ��������Ŵ�ѧ����ä���з���MCTP2����ͻ��
��ܿ, ����ΰ, Ingo Kennerknecht, ����, ֣���, ����ϼ������
��ҽ�ƴ�ѧ ��ҽѧ��ѧ�������ģ������Կ�ѧ�о����������廪������ѧ�о����ġ�����IDG/McGovern���о���������������ѧѧԺ��������ʵ���ҷ�������ѧ�о���������ѧԺ�˴Ź���������ġ�����ҽѧ�����빤���ص�ʵ���ң���������ѧԺ�������о������¹�Westfälische Wilhelms��ѧ�����Ŵ��о�����������ѧ������ѧѧԺ
ժҪ
��ʶ������Ӿ���֪�������֪������Ҫ����Ȼ��ä��prosopagnosia��face blindness���Ѿ�����������ʮ���꣬�����췴Ӧ��Ԫ�ѱ����ֽ�������ͣ�������Ŵ�ѧ������������ڴ����DZ����þ����Ŵ�ѧ���ִ�������ѧ�о�17��Ľ������һ����18λ���ճ�������ʶ�����ѵ�������ä��congenital prosopagnosia, CP����Ա�ļ�ϵ��������MCTP2�����ҵ���ȫ������Ķ���ͻ�䣬���������ڱ����һ���ƽ�ϵĿ�Ĥ���ס��ڸ���CP���塢�Լ�6589����ɸ�鵽��CP�з��ָ���MCTP2 �ĺ���ͻ�䡣14λMCTP2��������ͻ��S80fs ��Я���ߣ���19λ��Я��������ʶ����������𡣰���һ����ͥ��10λ��Ա��6����ͥ����ʾS80fs��CP������ԡ����ܺ˴Ź��������MCTP2ͻ���CP�Ը�����ʶ����ϰ�����������Ҳ�Ĵ�����ʶ��������ظ������ظ�ѹ�Ƽ��͡����緢����ä��76���һ����ä���˷���47������ǵĽ����ʾ��MCTP2 ͻ����CP�Ļ�����ԡ����ǵ�һ�η���������Ӿ������֪������Ļ���
����
��Ȼ������֪�dz���Ȥ����������֪�ķ����о�ȴ������ͨ�������ڶ����о�ijЩ����Ȼ���������о����ǡ�����;���������о��ڶ�������еı��ͣ����ҳ����ǵ͵ȶ����Ϊ�ڷ����鳤�ද�����ͨ�������о����ֻ�����ˣ�����ֻ��������ڵ���֪�����ڵ͵ȶ��ﲻ���ڵ���֪���з����о���ԶԶ����ڶ��ڼ���Ϊ�ķ����о���
���ǣ������༲�����Ŵ��о��dz��ɹ���Gusella et al., 1983; Schrott et al., 1972; Tsui et al., 1985���� �������Ŵ�ѧ�о��Ŀ�������˵��û�б�Ҫ�ٶ�������֪�뼲���и����IJ���Ŵ�ѧ�ṩ�˿����о�������֪����;������ʮ����������ʵ���ҽ����˼���������֪��ȫ����������о���GWAS�����Ӽ��䡢�������ԡ����Ӿ���֪�ĸ�֪�л������п��ƣ�Chen et al., 2018a; Chen et al., 2018b; Zhu et al., 2019; Zhu et al., 2016; Zhu et al., 2018; Zhu et al., 2020������Ȼ���Ƿ�������صı�ǣ������Dz�֪�����DZ�ǵĻ����Ǹ����Ļ��������о�����֪��û�������ϵ���Ŵ�ѧ�ϣ����ϵ�����������ɹ����ҵ����༲���Ļ���ͻ�䡣�����ô�;���о���ʶ��ķ����Ŵ��������о�����ڴˡ�
��ʶ���Ӿ�ʶ�����֮һ��Ϊ�����֪�����裬ͨ�����������ڸߵȶ��Bruce and Young, 1986; Freiwald et al., 2016; Grill-Spector and Malach, 2004; Tsao and Livingstone, 2008���� ��������������ں�����ҶƤ�㣨IT����Desimone et al., 1984; Gross et al., 1972; Rolls, 1984; Yamane et al., 1988������Ϲ���STS����Baylis et al., 1985; Bruce et al., 1981; Chitty et al., 1985; Perrett et al., 1987; Perrett et al., 1982; Perrett et al., 1984, 1985; Rolls, 1984; Rolls et al., 1985; Saito et al., 1986�������˶��������췴Ӧ����Ԫ����̼����Ե�ʵ����֧������Ԫ����ʶ��Ĺ������壨Afraz et al., 2006; Moeller et al., 2017������Ԫ�������췴ӦҲ���˵��¼���ص�λ�У�Allison et al., 1999; McCarthy et al., 1999; Puce et al., 1999����ֱ�ӵ��¼��Kreiman et al., 2000��������Ҷ���Ҷ�������췴Ӧ������Ҳͨ��������ɨ�裨PET����Haxby et al., 1994; Sergent et al., 1992������ʹŹ������fMRI����Barraclough and Perrett, 2011; Clark et al., 1996; Dzhelyova et al., 2011; Kanwisher et al., 1997; Liu et al., 2010; Malach et al., 1995; McCarthy et al., 1997; Puce et al., 1996; Tsao et al., 2003; Tsao et al., 2006; Tsao et al., 2008���������м������Ŵ̼��Ľ��Ҳ����Щ�������ʶ������Ҫ�Ŀ�����һ�£�Pitcher et al., 2009; Pitcher et al., 2008; Polk et al., 2007����
��ä��prosopagnosia��, ����Ϊ�¹����ڿ�ҽ��Joachim Bodamer��������Bodamer, 1947��, ����ʶ���ȱ�ݣ��������ڵͲ��Ӿ���߲��������⡣����Ϊֹ���ڷ���ˮƽ������ä����ʶ��һ����֪��
������ä��congenital prosopagnosia, CP��, Ҳ��Ϊ�����Ի��Ŵ�����ä��OMIM 610382��, �����1976�걻������McConachie, 1976���Ƕ������Ӿ�ѧϰ��ʶ���ѡ������ȱ���κο��Լ��������ˣ�Behrmann and Avidan, 2005; Damasio et al., 1990; Duchaine and Nakayama, 2006b; Gruter et al., 2008; Kress and Daum, 2003; McConachie, 1976; Nunn et al., 2001; Susilo and Duchaine, 2013�����ʾ�ʽ��ɸѡ������Kennerknecht, 2021; Kennerknecht et al., 2006; Kennerknecht et al., 2008a; Kennerknecht et al., 2007������Ϊ���ԣ�Bowles et al., 2009��������һ����ȺCP��������1.8 to 2.9%, ȫ�������ǧ��CP���塣��ϵ�о���De Haan, 1999; Dobel et al., 2007; Duchaine et al., 2007; Galaburda and Duchaine, 2003; Grueter et al., 2007; Johnen et al., 2014; Kennerknecht et al., 2006; Kennerknecht et al., 2008b; Kennerknecht et al., 2007; Lee et al., 2010; McConachie, 1976; Schmalzl et al., 2008����˫�����о���McKone and Palermo, 2010; Polk et al., 2007; Wilmer et al., 2010; Zhu et al., 2010����ʾCP����ʶ���������и߶��Ŵ��ԡ��������۲쵽�ij�Ⱦɫ�������Ŵ�ģʽ��De Haan, 1999; Duchaine et al., 2007; Grueter et al., 2007; Kennerknecht et al., 2006; Lee et al., 2010; Schmalzl et al., 2008��, ��ʾ���������ͻ����Ե�����ʶ��ȱ�ݡ�
���Ǵ�һ����18λ�ճ�������ʶ�������ѵĴ��ϵ������������һ��CP ������롰���� C2 �����Ĥ2�������ʣ�MCTP2, GenBank��NM_018349�������ճ�������ʶ�����ѵ������ҵ������MCTP2 ����ͻ�������ԡ�fMRI�����ʾMCTP2ͻ��CP���߶Ը�����ʶ��ȱ�� ������Ҳ�Ĵ���������rFFA����ͬ�������ظ����ַ�Ӧ�쳣�����ǵķ��ֿ��Դ̼����Ŵ�ѧ�ͻ�����ѧ�о��������֪��
���
������CP���ߵļ�ϵ
������һ��������ͥ����ͥA����35λ��Ա����1, ����S1, ͼ1a����18��ͥ��Ա�������ճ�����������ʶ�����ѣ�9λ��IV:2, IV:4, V:9, V:10, V:11, V15, VI:5, VI:6 and VI:9����������ϵ֮ǰ��֪�������������������һ���⣻4λ��IV:6, V:4, V:6 and VI:8���ܳ�ʱ�������п��ܵ���֪��ԭ��; 5 λ��IV:10, V:1, V:13, V:19 and VI:7���������ճ���������һ���⣬����ȡ��ʩ������֮�������IJ��ԣ���1�����Զ�ע�ӽӴ���������ʶ�����ѻ��ô�������֮���������ĸ���أ�p < 0.001, r = -0.658, n = 35����
����ǰ�о���Johnen et al., 2014; Schmalzl et al., 2008��һ��, ��ͥ�ھ��������ԣ������ǽ�����������-���ģ�Cambridge Face Memory Test-Chinese�� CFMT-C����Bowles et al., 2009; McKone et al., 2012��, ��Խ��ų������⣨the matched Cambridge Car Memory Test��CCMT����Dennett et al., 2012������������ЧӦ��the Face Inversion Effect��FIE�������⣨Yovel and Kanwisher, 2005����14λ����ʵ��������ʶ�����ѵļ�ͥ��Ա��10λ������һ����Ϊ��������ȱ�ݡ�V:1, V:9, V:11, V:13, V:15, V:19��VI:8��CFMT-C�÷�����һ��������������ˮƽ���������SDs��, ��V:4��z = -1.60����V:6��z = -1.61����ΪҲ���Բ����-1.5 SDs�����ڼ������ӹ���FIE��������, V:6��z = -1.62��, V:9��z = -1.62��, V:11��z = -2.56����VI:5��z = -1.81���ĵ���ЧӦȱ�ݡ�V:1��z = -2.96��, V:13��z = -1.73��, V:15��z = -1.55����V:19��z = -1.52������������������Ҳ�ܲ����λ��V:10, VI:6, VI:7��VI:9���ճ������ѣ�������ʶ��ļ���б������졣���Ƕ���CCMT���ܺã���ʾ����ʶ�����������̼�����������������ʶ�����ء�ÿλ��Ա��Z�ּ���1��
Ⱦɫ��15q����������ѡCP�����
Ϊ�˴��ڹ���С�ģ���ͥA�ij���������������Ϊ����1����������������S1�����������9����ͥ��ԱΪCP���ߣ�V:1, V:4, V:6, V:9, V:11, V:13, V:15, V:19 and VI:8������ϱ���������̸��ʾ����ʶ���쳣�����һ���CFMT��Ϊ�����ֲ���LOD��logarithm of odds������Ϊ3.49, ��ʾ������һ��CP��������һ��3.9����ԣ�Mb������Ҫ��ѡ����major candidate region��MCR����rs12148885��chr15:94251330, 15q26.1���絽rs288435��chr15:98160439, 15q26.2������Ӧ��15��Ⱦɫ����1-LOD �½���, ������hg19��װ��ͼ1b����������ɸѡ�����ʾ�ڸ�ͼS3a��һǧ���ظ��ļ����ģ����ʾ����pΪ0.023, ��ʾ��һ����ڻ�����ˮƽ�������ԡ���һ����9q21.13 ��LOD��������1.0����ͼS3a��, �������ģ��û�дﵽ�����������ԡ�MCR��ͬ���ĵ����ͼ���ͼS4a��
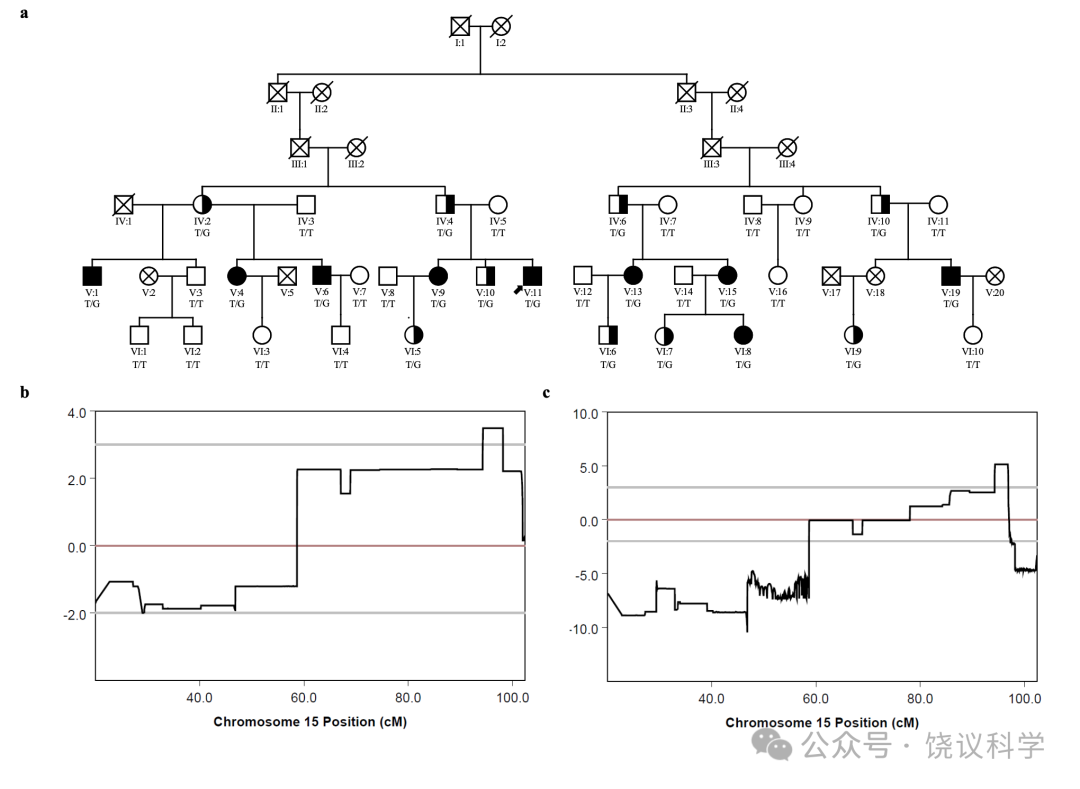
�п�����Щ������Ӧ�Բ�������˸���ʶ�𡢶�������ʶ�𣬵��·����ģ����Dalrymple and Palermo, 2016; Duchaine and Weidenfeld, 2003; Grueter et al., 2007������ˣ����ǽ����˵ڶ�����������������2������������������1�ڻ�����������4λ�ճ�����ʶ�����������Ϊ�����������ģ�V:10, VI:5, VI:6 and VI:7����4λ�ճ���ʶ���������֤�ߣ�IV:2, IV:4, IV:6 and IV:10������S1��������2��������������1������֧��rs6497114��chr15:94245722����rs11045��chr15��96883321��֮�������LOD��������3�����LOD����= 5.13��, ��Ⱦɫ��15q26.1��26.2֮���2.64 Mb��ͼ1c, ��ͼS3b������һ����10q24.2��LOD ��������1��ģ���15��Ⱦɫ�������LOD������ͳ�������ԣ�p< 0.008����MCR��ͬ���ĵ����ͼ���ͼS4b��
MCR��������NCBI���Ͽ⣨https://www.ncbi.nlm.nih.gov/genome/gdv/��ע�͵ĺ�ѡ�������ڸ���S7��
ȫ����������ʾMCTP2 �����һ��ͻ��
Ϊ���ҵ�A��ͥ����CP��Ⱦɫ��15q26.1-26.2�����Ӱ����ʶ��������ϵͻ�䣬���Ƕ�����1��9λ��Ӱ����壨V:1, V:4, V:6, V:9, V:11, V:13, V:15, V:19��VI:8, ��S1������ȫ���������WGS����MCR��, Ⱦɫ��chr15:94983466��MCTP2ֻ��һ�����죨NM_018349.4:c.2147T>G, NP_060819.3:��p.I716S������ȫ��9λCP�����Ӻ��塣�������ͻ�䣨c.2147T>G��ΪA��ͥ���У���������dbSNP��v150��, ExAC03��genomAD��v2.1.1���������Ͽ⣬��Ȼ��һλ�û�������������λ����������ԣ�SNP��λ��rs200314451 ���ڶ���gnomAD v2.1.1�⣬NM_018349.4:c.2147T>C, NP_060819.3:p.I716T����С��λ����Ƶ�ʣ�MAF��0.000���ڣ�NM_018349.4:c.2147T>A, NP_060819.3:p.I716N��MAF 0.0001088���ڡ������������3600 �й��˶����У�����û�м�ͬһͻ�䡣
Ϊ���о���һ�����Ƿ��CP��ǿ��ѡ�ߣ������ڼ�ϵֱ�Ӽ�ϵSanger�������й����������������������1��9λ, MCTP2�� c.2147T>G [p.I716S]Ҳ������4λ��ʶ��������������Ϊ���䲻��������Ϊ����ǰ����Ҳ������CFMT-C����������ճ�������ʶ������������5λ��ͥ��Ա��ͼ1a, ��ʵ���ţ���
����, MCTP2 ����NP_060819.3:p.I716S��ͻ�俴������һ��CP��ϵ�������ճ���ʶ������IJ�����MCRΨһ�����Ĺ��ܱ��졣SIFT��Ng and Henikoff, 2001��Ԥ��I716S�²���MCTP2����ȫ��ռ������DNA��180ǧ�����, ��22�������������, ����ĵ�������878 ��������л����ֳ�����C2�����������Ĥ����TMRs����ͼS5a������һ��TMR��NP_060819.3:I716�ڷ�չ���ܺ����ӹ����鳤��߶ȱ��أ���ͼS5a��S5b����Freiwald et al., 2016; Hung et al., 2015; Moeller et al., 2008; Tsao et al., 2003; Tsao et al., 2006; Tsao et al., 2008����
����ɸѡ�ҵ�����CP��CP��ϵ������MCTP2 ͻ��
�����ۺ�ʵ�鿼�ǵĻ����ϣ������Ĺ��ܵ�λ������Ϊ�DZ����Ŵ�����Ҫ�����ߣ�Cirulli and Goldstein, 2010; Heinzen et al., 2015; Jordan et al., 2010; Pritchard, 2001�����ӵ��͵�A��ͥ��õĽ����ʾ�������������е�MCTP2������ͻ�������CP�����йء�
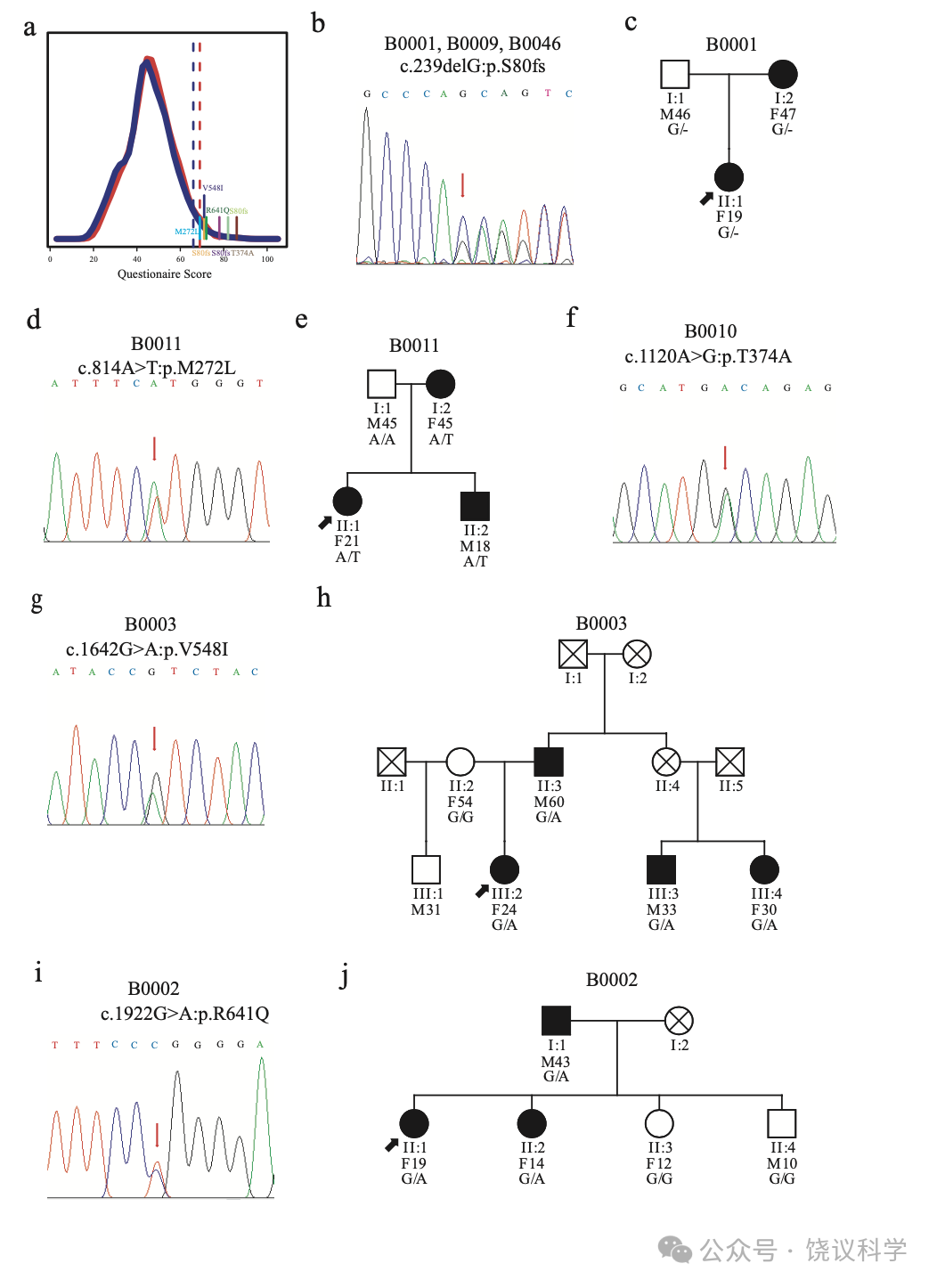
���ʾ�ɸѡ�ĵ�һ�����е�2904�ˣ�����������, �����ҵ�75λ������ƽ�����ٲ�2 SDs��������MCTP2 �����ӵ����С�7λ�з�����5�������Ӻ��ӹ��ܱ����壬����һ������ͻ���4������ͻ�䣨ͼ2a��ͼS5a, ����S8����
MCTP2����������1������ͻ�䣨NM_018349.4:c.239delG, NP_060819.3:p.S80fs�����ڱ�����ʶ�����ѵ�75λ�е�3λ������B0001, B0009��B0046����ͼ2b������һ�����Խ�ʾ����3λЯ��p.S80fs�ı��泤�������˵���ʶ�����۲�ȷ���ԣ�����������ʶ����һ���⣨����S2��B0001��һλ����Я��p.S80fs����ȡ�˹۲����������IJ��ԣ���������Ҫ����ʱ��ʶ���ˣ�ͼ2c, ����S2����
�ڸ���B0011�ļ�ͥ, ��������5��NP_060819.3:p.M272Lͻ�����Щ�˱��������Լ����˵IJ��ԣ�����������Ӧ�������ѣ�ͼ2d��2e, ��S2����������8��NP_060819.3:p.T374Aͻ�����B0010���壬�Լ���ʶ�������ص���ʶ��ȱ�ݣ�ͼ2f, ����S2���� B0003�ļ�ͥ, ������12��NP_060819.3:p.V548Iͻ�����������Ժܺã�ͼ2g��2h, ����S2����B0002�ļ�ͥ, NP_060819.3:p.R641Qͻ�����ͥ�����б��Ͷ�ͯ�����루ͼ2i��2j������Щͻ��λ���ڶ�������б��أ�ͼS5b����
���ݻ������ط�����ʾMCTP2������λ��������ʶ��Ĺ���
��һ���������ڵڶ�����1928�˵Ķ�����Ҳ���ʾ���飬�õ���ʶ����������������MCTP2��λ����Ե����ʱ������жȻ��ض�Ӱ��ĺ������������ԡ�
���ڸ��ɲ��飬�����Զ����У���ʹ���У�������ƻ��Ա����ȫ������ͻ����������أ����б���pburden = 0.0009, poptical = 0.0021���������к���ͻ��pburden = 0.0032, poptical = 0.0063����Ů�Զ��к�ͬ�������û�й۲쵽����ԡ���2��ʾ��������Щ�����Լ��ı����ϸ�ڡ����ǵĽ����ʾMCTP2����߱��������������ͬһ���������ã���ʾ���Զ��б��츺������ʶ�����������ٲ�������Ϊ��MCTP2���á�
CP����������ͻ��
��ϵ��Ⱥ���о����֧�ָı䵰���ʱ����MCTP2������λ��������ʶ��������أ���ָMCTP2������ʶ������������2904λ����ɸѡ��75λ��ʶ��������3λЯ��MCTP2�����һ�������ӵ�����ͻ��c.239delG��p.S80fs������ȥ��MCTP2�����ʵĴ֡�������Ƶ�����ѱ�����CP�����ʷ�Χ�У��㹻�ߵ�����ͨ���������;����ͬ�������ʶ��������Ҹ����Я���ߡ�
���ǽ�һ���ڵ�������1757�˵Ķ�����������һ����ͻ�䡣���Ǽ�16λ�Ӻ���Я����MAFΪ0.0046������14��ͬ���һ����飨����S3�������ǵ����Լ�¼��14λЯ������19λ����ͬһ���еķ�Я��־Ը��֮���˲���ʶ����Ϊ�Ķ�����𣨱�3����
��λ��C2180��C2666��C3164��C3282����������ϵ֮ǰ����ʶ������ʶ�����ѡ����C2666��C3164��ȫ���ܹ����ļ�ͥ��Ա��ʾ��һͻ������������ͥ������ʶ��ȱ�ݹ����루ͼ3a��3b, ����S3����
C3049û�б����Լ����ˡ������п��ܣ����Ǹе��Լ������������ˡ�������ͬ������S3����C3049�ļ�ͥ��ͼ3c, ����S3����, ͻ�仹�����������ݣ�II:2 and III:1��, �����ճ��������ѡ�
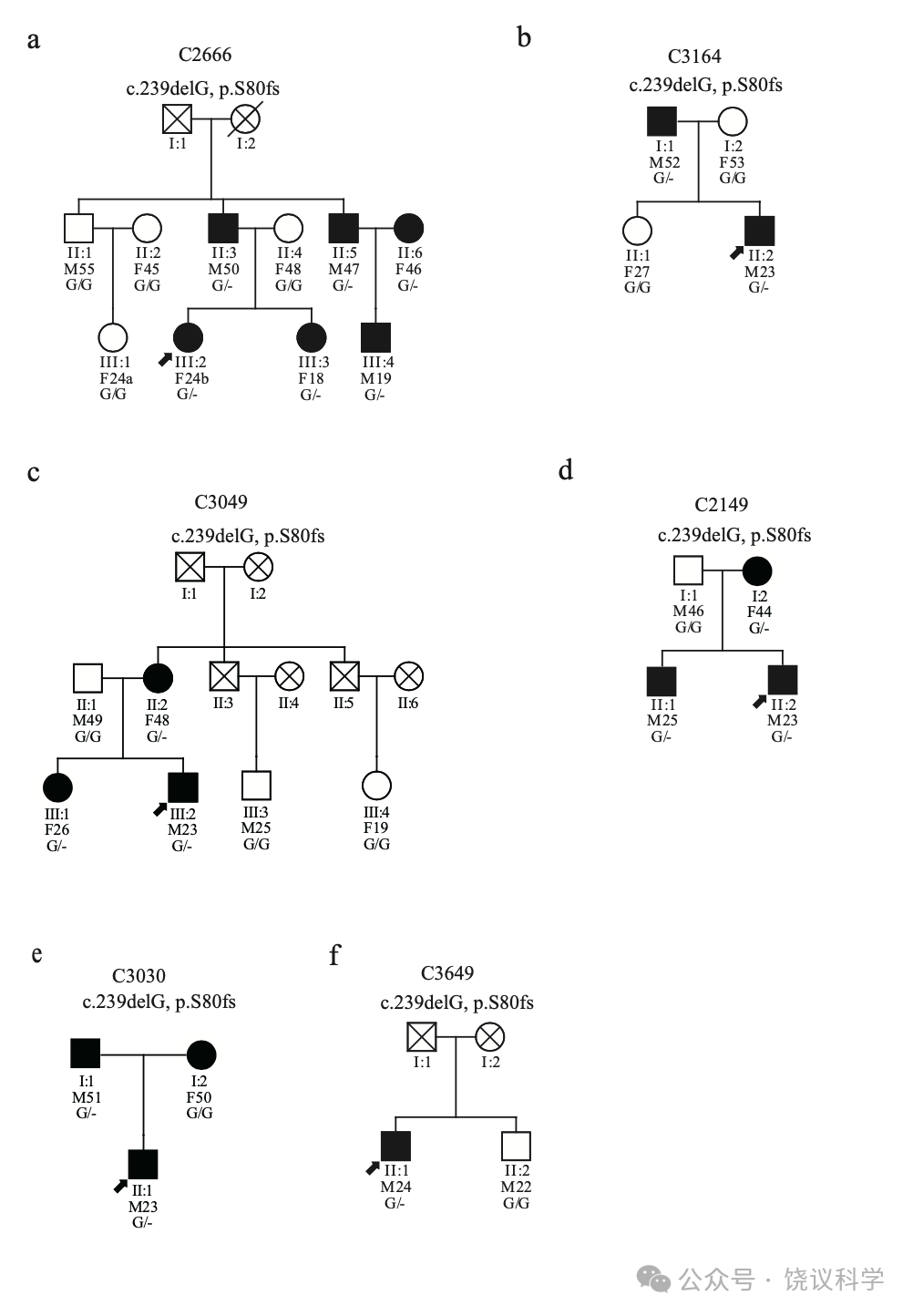
����9λ����������Ϊ�Լ���������һ����������������������S3�������ǣ�����8λ��C2149��C2259��C3030��C3234��C3358��C3420��C3649 ��C3731����չ����Ӧ����Ϊ��������ȷ��ѧϰ���������������ճ�ʶ�����ˣ�ȫ��, �������������ѡ�����������û�����������ˣ���Ļ�ϵ���Ա���ر���Ů��Ա�����������˵����볡�������ԾͲ��ᶼ�ܺõع������������ٵ��ڡ����ܿ������Ϣ��C2149��Я����һͻ������ݱ�Թʶ�����ѣ�ͼ3d, ����S3����C3030�����˵IJ����Ŵ��Ը�ĸ��ͼ3e, ����S3����C3649������û����ä��Ҳ��Я����һͻ�䣨ͼ3f, ����S3����C2147������������û����ʾ���Ե��쳣��ʶ��
ͨ���ռ�ĸ-��-������˵��������ϣ���ͼS6�����ų���ͬ������Դ�Ŀ����ԣ����Ƴ�c.239delG��p.S80fs��ͻ�����Ʋ�ĵ����͡�
MCTP2 ͻ��CP���������о�
��A�����6λ��Ա�����Ƿ���FFA����������쳣��Ӧ��MCTP2 �����I716Sͻ����ء�
�������о�һֱ�۲쵽rFFA���������Ĵ��ں����ֲ�ͬ������Grill-Spector et al., 2004; Haxby et al., 2000; Kanwisher et al., 1997; Puce et al., 1996; Rangarajan et al., 2014; Rotshtein et al., 2005; Yovel and Kanwisher, 2004; Zhang et al., 2012�������������������ߵ�rFFAs����ͼS7a��, ��I716Sͻ�䣨A-VI:5, ��ͼS7b, A-VI:7, ��ͼS7c��A-VI:9, ��ͼ S7d����û��I716Sͻ�䣨A-VI:1, ��ͼS7e, A-VI:2, ��ͼS7f ��AVI:10, ��ͼS7g���ļ�ͥ��Ա����������rFFAs�����Щ��ʾ��������ͻ�䣬A��ͥ��Ա��rFFA������ض����з�Ӧ��
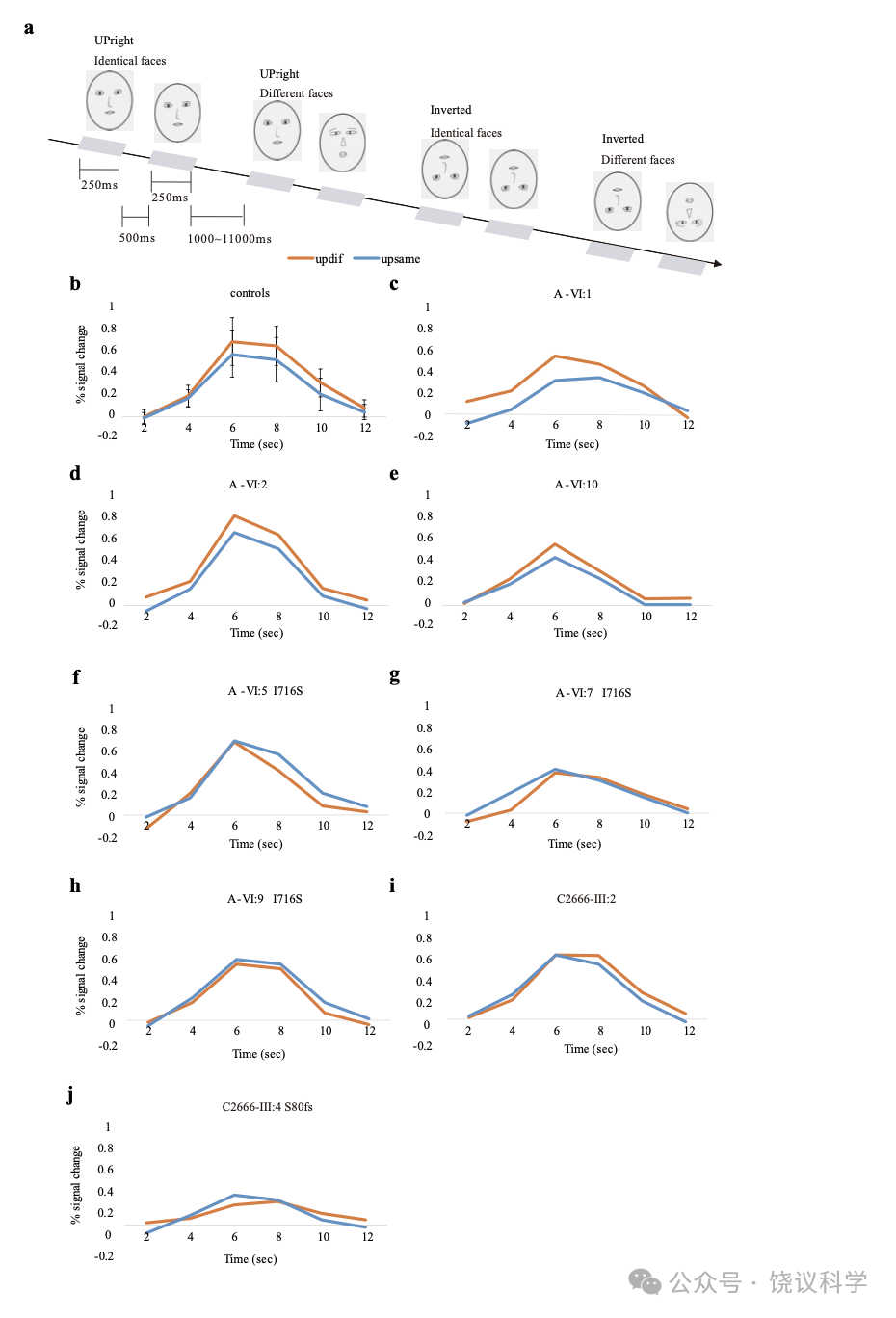
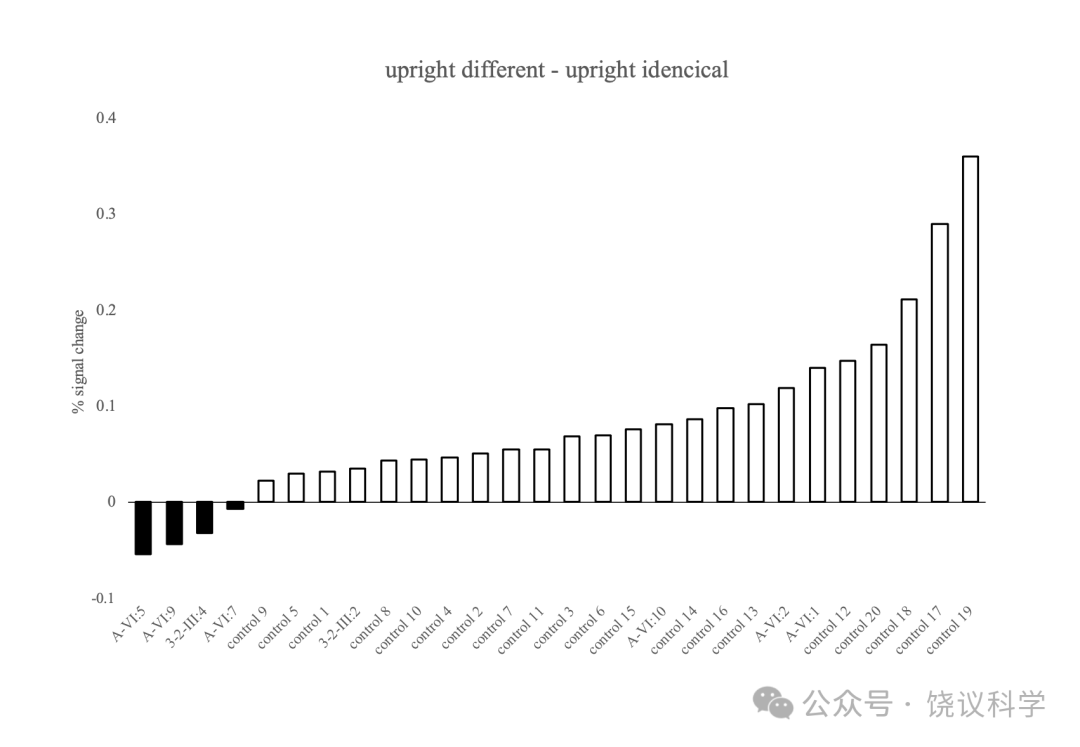
Ϊ�˼���ͥ��Ա��rFFA�ܷ�ӹ����ĸ�����죬�������¼�������fMRI��Ӧģʽ����ʵ�飨Yovel and Kanwisher, 2005����ͼ4a�����������飬fMRI��������������Ӧ�Ŀ�Ԥ�Ƶ��ظ�ѹ����rFFA�߶���������� t-���� p < 0.0001, ͼ4b�������ң�ÿ���������߶�һ�����õIJ�ͬ������UDF��������Ļˮƽ������һ�����õ���ͬ������UIF������UDF>UIF�� �IJ����20λ�����˵�18���г�p < 0.05��������������������������Ԥ�Ƶķ����з����������ƣ�p < 0.057, p < 0.25������ÿһλû��I716Sͻ��ļ�ͥ��Ա��A-VI:1, ͼ4c, A-VI:2, ͼ4d ��A-VI:10, ͼ4e����UDF������Ļ����UIF�ԡ���UDF>UIF�������ÿһλû��MCTP2ͻ��ļ�ͥ��Ա��p < 0.05�������ԡ��෴����I716Sͻ��ļ�ͥ��Ա��A-VI:5, ͼ4f, A-VI:7, ͼ4g and A-VI:9,ͼ4h��֮rFFA����fMRI�źŶԲ�ͬ���ķ�Ӧ����������ͬ������I716Sͻ��ļ�ͥ��Ա����Ӧ�֣��ڸ���Ȥ����ROI�����ò�ͬ������������ͬ�����ٷֱȵı仯������ÿһλ�������������ߣ�ͼ5������I716Sͻ��ļ�ͥ��Ա�Ա�������������ͬ����ȱ�����������ƣ�A-VI:5, p = 0.05407, A-VI:7, p = 0.12832 ��A-VI:9, p = 0.06548������Ϊ�ظ�ͬ����������rFFA����Ϳ�����Ҳ����I716Sͻ��ĸ�����٣�����ͬ����CP����CP��ͥ��Ա�����������������Ƶ�rFFA�����Щ�����ʾMCTP2����ͻ��ļ�ͥ��ԱrFFA����ͬ���ķ�Ӧ�������������л��ƽ���Ϊʲôʶ���Ѿ��������������ѡ�
C2666����MCTP2�����c.239delG��p.S80fs��ͻ��, ���ǹ۲쵽p.S80fs��III:4����ͼS7h������p.S80fs��III:2����ͼS7i����rFFA��������������Ӧ�������ճ���ʶ�����ѵĸ���III:4��ͼ4j����rFFA֮fMRI��ͬ�������ظ�Ҳȱ����Ӧ��
����
��ʶ��ķ����Ŵ�ѧ�о�
��äΪ������֪����ʶ�𣩵��Ŵ�����ģ�͡��Ѿ��������������CP��ϵ��Duchaine et al., 2007; Johnen et al., 2014; Schmalzl et al., 2008��, �����²�������������ǵ��о�������˵�Ƕ�������֪�ĵ�һ�������Ŵ�ѧ�о���������˵���������ͥ����ʶ��ĵ�һ������Ŵ�ѧ�о���
���ǵķ���MCTP2�����ͻ�������CP�й��ס���һ����������һ�£�1������������Ⱦɫ������CP��ͥA����λ��15q26.1-q26.2��CPλ�㣨ͼ1, ��ͼS3��; 2��һ��MCTP2 ���������ͻ��c.2147T>G��p.I716S����A��ϵ����MCRȫ����������ҵ���Ψһ��ȫ��CP������Ŀ��Ըı䵰�������е�ͻ�䣨ͼ1��; 3����2904����ɵĶ������ҵ���75λ��ʶ�����ѵĸ�����7λ��ä���з���MCTP2 ����5������ͻ�䣨Figure 2��; 4����7λ��ä���У�4λ�ļ����ԱԸ������������ʾ����������͵�����ԣ�ͼ2c, 2e, 2h��2j��; 5��1757�����ߵ���һ������16λЯ�����4������3λ��ä������ͬ����ȱʧͻ��c.239delG��p.S80fs��, ����14Ը����һ��������14λЯ�����ճ���ʶ����Ϊ��ͬ������ͬһ���е�19λ��Я���ߣ�ͼ3��; 6��14λ��4λ�����ɵ���ä������������ϵԸ�������������֧��c.239delG��p.S80fs������ä��أ�ͼ3a��3b��, ��һ����֧�����Ը����ϵ��ͼ3c��3d��3e��3f�������Ƿ�չ����ȷ�IJ����÷����������˷�����ʶ������; 7����1928λ��ɵĶ��л�����ط���Ҳ����MCTP2������λ��������ʶ������������ԣ�8���������о��У�MCTP2����ͻ��ļ����Ա��ʶ��ȱ����rFFA�Ը�������Ӧ�쳣�������
CP�����
��ͬ����֪�����о�CP��ʾ�����ԣ�Behrmann and Avidan, 2005; Kress and Daum, 2003; Le Grand et al., 2006; White and Burton, 2022������Ȼ�����ⱻ֤�����ã�Duchaine and Nakayama, 2006a��, û��һ�������ļ�����㹻����ȫ��CP��������Duchaine and Nakayama, 2006a; Duchaine and Nakayama, 2004; Duchaine and Weidenfeld, 2003; Grueter et al., 2007; Shah et al., 2015����
�������ĸ��ض�������ʹ��ͬһ����ͥ���Ƶ��Ŵ��ͻ��������У�Johnen et al., 2014; Lee et al., 2010; Schmalzl et al., 2008��������Ҫ������Ϊ������CP���������ܺܶ�����Ӱ�죬������Ϊ������������֪���ܣ������ⷶʽ�ľ���������ЩCP����������ʶ���������ѣ�����������Ϊ�����ȴ���ܱ����൱�á����ӹ�������ֲ����Ԥ����ʶ��ȱ�ݣ����෴��������
��������Ϊ��������ʶ��Ҳ������Ҫ�����������ұ��档������Ҫ����Dunning-Kruger��DK��ЧӦ��һ��Ԫ��֪����, ��ij��������ֲ���˴��������Ϊ�������˸��ã������ֺܺõ�������Ϊ���������ˣ�Dunning et al., 2003; Kruger and Dunning, 1999����DKЧӦ������ʶ�����ұ��棨���ҹ��ƣ������˹��ƣ�ͬ�й��ƣ���Zhou and Jenkins, 2020����������ʶ�����ұ�����ʱ��Ҫ�������ܿ��ܲ�����֪������ʶ��ȱ�ݡ�����ͨ����ϸ���ԣ����Է��������в��Ժ��ճ�ϰ�ߡ�ͬʱ����DKЧӦ�У���Ȼ�õ������ߵ�������������ֻ�����Լ��൱��ƽ��������ⲻ������Ӱ���������жϣ��������ǻ�������Ϊ��������ˣ������ǵ��о��У�����ͨ����ϸ���Լ�ϵ��Ա��Ϊ��Ҫ����ϻ�����
MCTP2������
MCTP2�������ĵ�������3��C2�����������Ĥ�������Ʋ���ͻ�����ݵĵ����ʣ�Shin et al., 2005������C2������Ca2+��Shin et al., 2005�����ܶ���Ca2+���C2 ����ĵ����ʲ���Ĥ������ת��, ��������ؼ����ã�Cho and Stahelin, 2006; Shupliakov and Brodin, 2010����
MCTP�൰���ʵ�C2�Ϳ�Ĥ���ڷǼ�����ͱ��أ����߳��Ӭֻ��һ��MCTP����, ������������������MCTP1 ��MCTP2����MCTP1������ֽ����ϵͳ������ָ����������ϵͳ��Ԫ��ͻ��������ѭ����Qiu et al., 2015�����ڹ�Ӭ�� MCTP�����ȶ�ͻ�����ݺ�����̬�����ԣ�Genc et al., 2017�����������4��MCTP����������ͼ���ϵͳ�� �õ�MCTP2b iӰ����̥������Espino-Saldaña et al., 2020����
�˵�MCTP2��MCTP1 ������߶��Ŵ�������ע��ȱ��/�ද֢��ADHD���й�����Kweon et al., 2018; Mick et al., 2010���� �������ӽ�ϡ��Ļ��������14��ADHD��ػ����и�����Poelmans et al., 2011���������������˫�����ҵ�MCTP1 ����������ԣ�SNP����Scott et al., 2009����Ҳ��һƪ�����������֢��MCTP2 SNP��Djurovic et al., 2009����
�������ᵽ��1757�˵ĵ����������У����ҵ�����Sanger������֤��MAF < 0.005��64λ��������λ�㺱�����졣��־Ըͬ��Ļ����Ͻ��������ԡ�16λ����Я�� ����ȷʵͻ�� c.239delG��p.S80fs����MAF of 0.0046����14λ���������ԡ����ص�S80fs����Я��������ʶ������ֱ��ά��������ԡ������ҵ�һ��Я��S80fs ��ͥ��C2666����10λ��Ա��ͻ������äҲ������ԡ���Ȥ���ǣ�����Я��S80fs ��II:5����ä��������II:6Ҳ��ͬ��ͻ�䲢����ʶ�����ѡ����ǽ�һ��������MCTP2����������Ƶͻ��ĸ��塣���и���ĸ��弰���ͥ������ͻ��ͼ���λ��ͻ��Ҳ��ʾ��ʶ���쳣��
�ӳ�ǧ�����õ��Ļ����������������ʾMCTP2�������������, �����Ҷ��McCall et al., 2011��������ѡ����һ�¿ɿ��ؼ��������FFA, ���ҶƤ��Ĵ��ص�һ��С�������Ե���ͼ����ʾ��MTCP2�����ʱ���ͷֲ������������ڷĴ��أ�Sjostedt et al., 2020����
fMRI��Ϊ����ڱ��͵ķ������ṩ�������Ի֮��Ļ�����ϵ
fMRI��ʾ�ض�����/��Ϊ����������Ԫ���������ڸ��ӵ�����Ϊ�쳣�������ڱ���������fMRI����Ϊ���Խ��ٴ���������õش�����������ѧ��Rasetti and Weinberger, 2011��������-�Ŵ������������ֲ�������ض�Σ�յ�λ������Ի��Ӱ�죬�������Ǽ���ϱ��͡�
�ڱ��о�����������Ӧ��ʽ��Grill-Spector et al., 2006; Grill-Spector and Malach, 2001; Henson and Rugg, 2003�������˳���-�����о�����ʾ��ͥ����ʶ�����ѳ�Ա��I716S��S80fsͻ���Ӱ�졣�����������ߡ��Լ���ͥ��û��MCTP2ͻ��ij�Ա�У����ظ�����ͬ��������ʱ��rFFA��������ź���ʾ���ͣ�����Я��MCTP2ͻ��ļ�ͥ��Ա��û��������Ӧ��
δ���о��У� fMRI���Կ���Ϊ��ʶ�����Ҫ�ڱ��͡�
������ʶ��Ļ���
���ǵ��Ŵ�����ṩ֤�ݱ���MCTP2����ͻ�䵼��������ä������fMRI�����ʾMCTP2�����������������Ҫ����·��
��Ȼ������֪��Ҫ�ܶ�ϸ���ͷ��ӡ�MCTP2������Ψһ����CP�Ļ���2904�˶����ҵ���75λCP������ֻ��7λЯ��MCTP2����ͻ�䡣��1928�˶�����Ů����ʶ��������MCTP2������λ����û������ԡ�����������о����а�����
���Ƿ��ָ����ͥû���Ŵ�����������Ҫ�������������Ļ�����������Կ������´���������������������ͺ����Ŵ����죬��Ҫ�����������������ˣ��ҵ������Ŵ�������Ҳ��֤�����й�����ʶ���MCTP2 �����
�����뷽��
�����ߺͷ���
������ϸ��
���о�Ϊ������ѧ��������Ͷ���������ίԱ����ͬ�⡣����ǰ���������䷨������������֪��ͬ������о������������о������������DNA��Gentra Puregene Blood Kit��QIAGEN����ȫѪwhole blood��BD Vacutainer 3.2% Sodium Citrate��1:9������á�����GeneFiX DNA Saliva Collector and Isolation Kit����Һ��á�
��ϵA
A��ϵ����֤��V:11��ͼ1a, ��1���������ҽ��ܡ��ñ��İ�ṹ�����ԣ�Carbon et al., 2007; Grueter et al., 2007; Kennerknecht, 2021; Kennerknecht et al., 2006�������˸ü�ϵ��35λ���壨18 CP��9��9λ������16��72��; 17λ��CP, 9��8Ů, ����15��78�꣩��25λ�������60�ļ���������˽�����������-���ģ�Cambridge Face Memory Test-Chinese�� CFMT-C����Bowles et al., 2009; McKone et al., 2012��, ��Խ��ų������⣨the matched Cambridge Car Memory Test��CCMT����Dennett et al., 2012����������ЧӦ��the Face Inversion Effect��FIE�������⣨Yovel and Kanwisher, 2005���� 34 individuals��not including VI:9, who contacted us voluntarily after we had finished genotyping��were included in the linkage study. Nine individuals��V:1, V:4, V:6, V:9, V:11, V:13, V:15, V:19 and VI:8��were selected for WGS. This information is listed in Table S1. None of the persons showed signs of an autism spectrum disorder or neurodegenerative disorders.
��һ��2904�˶�����ɸѡ���ĸ�����MCTP2 ͻ���CP����
To investigate more CP individuals and examine whether rare functional mutations inMCTP2 are present in more CPs, the first cohort of 2904 individuals��average age of 19.25 + 1.30, 2161 females, 743 males, Jiangxi province ��from the Westlake BioBank for Chinese��WBBC��pilot project��Zhu et al. 2021a; Cong et al. 2022a; Cong et al. 2022b��, were screened with a questionnaire adapted from the 20-item self-report measure��see CP Questionnaire in Methods Details��for quantifying CP traits. 78 individuals scored worse than the mean by 2 SDs��Figure 2a��. 75 of them provided DNA samples. 44 individuals including 7 individuals with MCTP2 mutations agreed to an interview for CP diagnosis and family members were contacted for availability for further studies.
�ڶ���1928�˶��еĻ��ڻ������ط���
The second cohort of 1928 individuals��average age of 18.51+ 0.93, 1085 females, 843 males, Jiangxi province��used in the gene based association of the rare functional alleles in MCTP2 with face recognition ability were also from the WBBC project��Zhu et al. 2021a; Cong et al. 2022a; Cong et al. 2022b��. The coding sequences of the MCTP2 gene for each person was analyzed by tagged-amplicon deep sequencing.
�ӵ�������1757�˶���ɸ����MCTP2����ͻ��c.239delG��p.S80fs������
A third cohort of 1757 individuals��average age of 19.13+ 1.07, 1295 females, 462 males, Guangdong province��were sequenced for the presence of the c.239delG��p.S80fs��mutation in the exons of MCTP2. The coding sequences of the MCTP2 gene for each person was analyzed by tagged-amplicon deep sequencing. For individuals carrying p.S80fs, their family members were further contacted to test for the S80fs mutation by direct Sanger sequencing and assessed by the standardized semi-structured interview. The differences in the behavior of daily face recognition between the 14 carriers and 19 non-carrying volunteers from the same cohort were analyzed by independent sample t-test.
��Ϊ���Ķ�������
338 normal participants��average age of 42.68+ 16.53, 164 females, 174 males��were tested. These participants were unrelated. Participants were not selected for face recognition ability with no known history of major brain injury, or other major disorders likely to affect face recognition��e.g., Alzheimer��s disease��, representing a random sample of the community. They were tested on a battery of tests including the CFMT-C, the CCMT, the FIE Test and the Cambridge Face Perception Test - Chinese��CFPT-C��.
fMRI������
Age-related dedifferentiation and compensatory changes in the functional network underlying face processing have been found in studies��Burianova et al., 2013; Germine et al., 2011; Goh et al., 2010; Le Grand et al., 2006; Lee et al., 2011; Park et al., 2012; Zebrowitz et al., 2016��. Therefore, in order to exclude age influence on fMRI results, we mainly studied family members younger than 30 years old. Family members who participated in the fMRI experiment were on a voluntary basis. In family A, 3 MCTP2 mutant CP individuals��VI:5, VI:7 and VI:9��and 3 non-mutant and non-CP individuals��VI:1, VI:2 and VI:10��took part in fMRI; in family 3-2, III:4��CP, with S80fs��and III:2��non-CP, without S80fs��took part in fMRI.
21 non-CP students��average age = 23.63+ 3.71, 6 females, 15 males��were recruited for fMRI analysis.
CP referred to those with obvious face recognition problems in daily life diagnosed by the structured interview.
All participants reported normal or corrected to normal vision, no history of neurological or psychiatric conditions and all were right-handed. Anatomical volumes��i.e., structural MRIs��had been routinely checked. One of the normal students was excluded from neuroimaging studies because the maximum head rotation was over 1.5 degree or the maximum translation was over 2 mm during localization.
We adopted the methods of studying single-cases that the severity of each individual could be reported by comparison with the control population��Bentin et al., 2007; Busigny and Rossion, 2010; Dricot et al., 2008; Gao et al., 2019; Hadjikhani and de Gelder, 2002; Righart et al., 2009; Rossion et al., 2011; Schiltz et al., 2006��. In fMRI analysis, each family member was compared individually to a small sample of normal controls by a modified t-test��Crawford and Garthwaite, 2002; Crawford et al., 2010; Crawford and Howell, 1998��.
����ϸ��
CP��ϵĽṹ����
The diagnosis of CP was based on a standardized semi-structured interview��Tables 1, S2 and S3����Grueter et al., 2007; Kennerknecht et al., 2006��,��һ��Grueter2008 which documented a variety of impressive qualitative differences between CP and non-CP, and had been validated with objective face recognition tests in previous studies��Carbon et al., 2007; Grueter et al., 2007��. These criteria were employed in recent literature on CP��Bate et al., 2019; Bate et al., 2014; Carbon et al., 2010; Dinkelacker et al., 2011; Esins et al., 2016; Esins et al., 2015; Gruter et al., 2009; Kennerknecht et al., 2008b; Stollhoff et al., 2010, 2011; Verfaillie et al., 2014; von Kriegstein et al., 2008; Zhao et al., 2018; Zhao et al., 2016��.
The interviewer asked questions in a semi-structured interview format with three or four questions about each diagnostic item. Interviews were held to embed the questions into conversations and questions about the same diagnostic items not asked sequentially. Interviews included a medical history in order to exclude conditions which might cause or mimic prosopagnosia. Affected people present a lack of confidence with face recognition. Symptoms include lasting and irritating subjective uncertainties of face recognition, failure to recognize familiar people out of context or in crowded places, overlooking familiar people and confusing strangers with familiar people, face recognition/learning time longer than socially accepted, development of adaptive behavior of critical situations and strategies for visual recognition of people and time of onset.
Consistent with the interview results, some individuals were aware of their CP before we contacted them. Some individuals who identify people via non-facial features such as voice, gait and general appearance and manner, or were unaware of face recognition problems, but had developed obvious compensatory strategies to cope with difficulties were also diagnosed as CPs.
CP�ʾ�
To effectively screen CP candidates from big samples, we adapted a Chinese 20-item version from the 20-item self-report measure for quantifying prosopagnosic traits��Shah et al., 2015��, which asks about tangible experiences. This 20-item Questionnaire was included in a set of questionnaires for many research purposes and filled out by two cohorts of individuals online. Invalid questionnaires were dropped due to no distinction between forward and reverse questions. 2904 valid questionnaires for the first cohort and 1928 valid questionnaires for the second cohort were collected. The internal reliability measured by Cronbach's �� was 0.828 and 0.902 respectively. In the first cohort, 343 individuals finished the questionnaire for a second time several weeks later and the correlation for the first and second results across each individual was very high by Pearson correlation coefficient analysis, r = 0.081, p < 0.001. There was a significant difference of the score distribution between the female and male participants��p < 0.001, two tailed t test��, so the candidate CPs were screened and the gene based association study were carried with different genders respectively. An individual with a score above 2 SDs of the mean of the controls was defined as a CP candidate.
��Ϊ���Ĵ̼��ͳ���
All behavioral tests were adapted and integrated into a whole set using the Hyper Text Markup Language. All participants were tested individually. The tests were run on a desktop PC with screen resolution 1024 �� 768, refresh rate 85Hz. Participants were seated at a viewing distance of approximately 50 cm from the screen. All participants were tested wearing their usual optical correction. Participants were asked to confirm that they could focus without seeing blur on the computer screen. No participant reported any difficulty with focus at these distances. Gray-scale adult Chinese faces were used.
������������-���ģ�CFMT-C��
The Chinese face version of CFMT��McKone et al., 2012��was kindly provided by Professor Jia Liu of Beijing Normal University which we integrated with other behavioral tests into a whole set, and performed according to the standard procedure including the practice phase, the ��Learn�� phase with 18 trials, the ��Novel Images�� phase with 30 trials and the final ��Novel Images with Noise�� with 24 trials. All faces were Chinese male, shown without hair or facial blemishes and with neutral expressions. Participants were instructed to press the key corresponding to the number 1, 2 and 3 below faces. The test includes a total of 72 trials. Scores were reported as percent correct across the full test.
���ų������⣨CCMT��
The CCMT is a test similar in the experimental design as CFMT, with stimuli replaced by whole cars��Dennett et al., 2012��. We used the CCMT as a control of the CFMT to test for potential general object recognition deficits and to quantify the individual ability of performing such kind of tests. The original version of CCMT was kindly provided by Professor Bradley Duchaine of Dartmouth College, USA, which we integrated with other behavioral tests into a whole set, and performed according to the standard procedure��Dennett et al., 2012��
��������֪���-���ģ�CFPT-C��
We developed a version of CFPT using Chinese faces as stimuli��CFPT-C��, according to the standard procedure��Duchaine et al., 2007��by morphing six different individuals with the target face, containing 88%, 76%, 64%, 52%, 40%, and 28% of the target face in turn. The Chinese faces were male, shown without hair or facial blemishes and with neutral expressions. All faces were photographs of Chinese students at Peking University, with written consent forms collected before photographing. Each individual was photographed in the same range of views and lighting conditions. Eight upright and eight inverted trials were intermixed, with the upright trial occurring first half the time. Participants had one minute to arrange six morphed faces according to their similarity to a target face by clicking on a face and indicating where that face should be moved by clicking in the area between two faces. Scores were computed according to the previous paper��Duchaine et al., 2007��. The internal reliability measured by Cronbach's �� in our sample of 170 individuals was 0.705 for upright faces, and 0.483 for inverted faces. However, after our pilot study, the CFPT-C was excluded from further study, because strategies such as just comparing partial facial features were used in normal people as in CPs and inconsistent results were reported by others��Anstey et al., 2005; Bowles et al., 2009; Duchaine et al., 2007��.
������ЧӦ��FIE�����ּ��
The stimuli consisted of 20 gray-scale individual face images, cropped using the same 4 X 4.5 cm oval window��4.6º X 5.2º of visual angle��to remove cues from the hairline and face contour. All faces were photographs of Chinese students at Peking University, with written consent forms collected before photographing. Photographs were not repeatedly used in different tests. Pairs of face stimuli were presented sequentially either upright or inverted in a randomized order. The first face stimulus was presented in the upper-left quadrant of the screen for 0.5 s. After an interstimulus interval of 0.5 s, the second stimulus was presented in the lower-right quadrant for 0.5 s. The next trial will not begin until a same or different response was made by pressing one of two keys to respond by the participant. 80 trials were conducted in this test, half of them with upright faces and half with inverted faces. In each condition��upright or inverted��, the chance with the two identical faces was 50%. A few practice trials were presented before the beginning of the experiment. Scores were reported as percent correct for each condition. For the measure of the FIE, the difference in performance level between upright and inverted faces, an FIE index was calculated by entering the correction of performance for upright and for inverted faces in the following formula��FIE =��upright �Cinverted��/��upright + inverted��.
��Ϊ�������Ϸ���
Due to possible effects of ageing and sex difference on the scores of the CFMT-C, CCMT, upright faces and inverted faces in the FIE and the FIE index, we used multiple stepwise regression analyses to identify the covariates��sex, age��specific to each trait from 338 participants��aged 15�C83 years, 164 females, 174 males��. The results of the CFMT-C and the CCMT showed noticeable age-related decline and sex differences��Figure S1��. The scores of upright faces and inverted faces in the FIE correlated with ageing but not sex��Figure S2��, while the FIE index was not affected by age or sex among the control samples.
Examining function curves suggested that the behavior performance remained stable across early middle age, but began to decline noticeably at approximately 50 years of age. This is consistent with previous studies. In terms of the validity of the behavioral scores, only individuals with ages under 60��average age = 36.51+ 12.34, 130 females, 138 males��were included to calculate the best estimate of CP cut-offs.
We used the fit-and-residual procedure to calculate the standard residual of each participant��Z score��for CP family members as described previously��Bowles et al., 2009��. The regression function describing the relationship between age, sex and CFMT was��CFMT score = -0.212 age + 6.188 sex + 73.724. The standard deviation��SD��of the controls�� residuals of CFMT score was 11.189. The regression function for CCMT was��CCMT score = -0.134 age - 2.968 sex + 69.444, and SD was 10.252. The regression function for upright faces was��Upright score = -0.170 age + 87.641, and SD was 8.862. Z score was calculated by dividing the participant��s residual by the SD of the controls�� residuals.
For the FIE index, there was no correlation with age or sex. Normalized Z score was calculated for each subject by subtracting the mean of the control sample and dividing by the control samples�� SD.
�Ŵ�����
����������ȫ������������
DNA samples were genotyped using Infinium Human OmniZhongHua BeadChips��Illumina��, and normalized bead intensity data obtained for each sample were converted into SNP genotypes using Genome Studio. SNPs were then selected according to the following parameters��genotyping rate > 95%, minor allelic frequency > 1% and no significant deviation from Hardy�CWeinberg proportions��p> 0.001��by PLINK software��Purcell et al., 2007��. Gender corresponding to each DNA sample was checked by analysis of X chromosome heterozygosity using PLINK. The initial Mendelian inheritance in family A was analyzed by PLINK and KING toolset��Manichaikul et al., 2010��.
�����о������Ϸ���
For the preliminary linkage analysis in Family A��named linkage 1����Table S1��, we diagnosed 9 family members as CP cases��V:1, V:4, V:6, V:9, V:11, V:13, V:15, V:19 and VI:8��with the criteria of not only abnormality in face recognition exhibited through interviews, but also poor with the CFMT behavior test��under -1.5 SDs��. 11 family members with normal daily face recognition by interview and Z scores of all the behavior tests over -1.5 SDs were taken as normal controls��V:3, V:7, V:8, V:12, V:14, V:16, VI:1, VI:2, VI:3, VI:4 and VI:10��. The remaining subjects in the pedigree were set as unknown at this stage of analysis, including those over 60 and not suitable for behavioral tests.
We did a second linkage analysis��linkage 2��after adding four individuals who had daily face recognition problems but appeared normal on behavioral tests��V:10, VI:5, VI:6 and VI:7��and four founders with daily face recognition problems��IV:2, IV:4, IV:6 and IV:10��as CPs��Table S1��, and the remaining six founders without daily face recognition problems��IV:3, IV:5, IV:7, IV:8, IV:9 and IV:11��as controls. They were set as unclear in the linkage analysis analyzed above��linkage 1��.
Parametric linkage analysis was performed with the Merlin programs��Abecasis et al. 2002��, assuming autosomal dominant inheritance with 100% penetrance, disease allele frequency 0.001, and phenocopy rate 0.05. Because the presence of linkage disequilibrium��LD��might inflate linkage statistics, LD maps were constructed with the PLINK tool within family members with the LD thresholds��r2 < 0.5��. Following data QC, 177,126��r2 < 0.5��informative SNPs were selected for linkage analysis with Merlin��Abecasis et al. 2002��. 1000 simulation analyses were performed to exclude false positive results due to random chances under the null hypothesis of no linkage, with simulated data, while maintaining the pedigree structure, allele frequencies, and recombination fraction. For positive regions, haplotypes were constructed and subsequently checked manually on the basis of the minimal number of recombination.
���������죨CNVs�����е�����
CNVs were identified for each member. Genotyping and signal intensity data were exported from the GenomeStudio software 2011.1��San Diego, CA, USA��. The subsequent CNV calling analyses were performed using PennCNV��v.2011 Jun16����Wang et al., 2007��according to the manual. No correlation was detected between the CNV genotypes and the phenotypes for each linkage analysis.
ȫ���������WGS��
WGS was performed by the Next-Generation Sequencing Center of Biomedical Pioneering Innovation Center, PKU. Sequencing libraries were built with NEBNext Ultra DNA Library Prep Kit for Illumina.150 bp paired-end sequencing was done on Illumina HiSeq 4000.
WGS�����Ϸ���
FastQC��http://www.bioinformatics.babraham.ac.uk/projects/fastqc/��was applied to perform quality checks on various aspects of sequencing quality. Low quality bases were removed by Trimmomatic software��Bolger et al. 2014��with the parameters HEADCROP:5, LEADING:25, TRAILING:25, SLIDINGWINDOW:4:15 MINLEN:35. After that, clean sequences were aligned to human genome build hg19 and SNPs, INDELs or structural variants were called by SPEEDSEQ��Chiang et al. 2015��. BAM files locally realigned around INDELs were used to determine the average coverage, using GATK DepthOfCoverage package and default settings��McKenna et al. 2010��. After the alignment and variant calling, the mean depth of all samples was 34, and 96% of the mapped bases were covered at more than 10-fold on average��Table S4��. In the linkage region, 100% coding bases were covered at more than 20-fold in at least one of the four individuals. All variants were annotated to RefSeq hg19 and ANNOVAR��Wang et al. 2010��were used to add alternative allele frequencies, variant effect predictions and functional annotations. We excluded variants with minor allele frequency��MAF��> 0.05 in multiple databases, including the dbSNP��v150����Sherry et al. 2001��, the 1000 Genomes Project��1000g2015aug����Genomes Project et al. 2010; Genomes Project et al. 2012; Genomes Project et al. 2015��, the Exome Aggregation Consortium��ExAC03����Lek et al. 2016��and the Genome Aggregation��gnomAD, v2.1.1, http://gnomad.broadinstitute.org��databases.
��ǷŴ��ӵ���Ȳ���
Target-specific primers forthe coding sequences and the exon-intron boundaries of the MCTP2 gene��GenBank��NM_018349��were designed with universal primer sequences��termed CS1 and CS2��appended at the 5��-end and sequencing was performed as described previously��Forshew et al. 2012��.
��ǷŴ�����Ȳ�������Ϸ���
The raw paired 150 bp-long reads were mapped to the human reference genome��build hg19��using BWA. GATK��McKenna et al. 2010��was then used to perform local realignment and recalibrate base quality scores, producing a BAM file for each individual. All variants were annotated to RefSeq hg19 and ten algorithms��Sift, Polyphen2_HDIV and HVAR, LRT, MutationTaster, MutationAssessor, FATHMM, PROVEAN, MutPred and VEST��integrated by ANNOVAR��Wang et al., 2010��were used to add alternative allele frequencies, variant effect predictions and functional annotations. Among these 10 algorithms, if a site was predicted as deleterious by Sift, LRT, FATHMM, or ROVEAN, as probably or possible damaging by Polyphen2HDIV or HVAR, as disease_causing_automatic or disease_causing by Mutation Taster, as high or medium by Mutation Assessor, and with scores greater than 0.5 by MutPred or VEST, it could be considered harmful for the specific algorithm. 91% of targeted bases were covered at �� 100-fold per individual��Table S4��. We manually inspected each mutation using the Integrative Genomics Viewer��Robinson et al. 2011; Thorvaldsdottir et al.2013��to rule out false positive findings. If the coverage for each exon was under 100-fold depth in each individual, we performed direct Sanger sequencing to verify these regions.
��1928����ɵĵڶ����ж�MCTP2���еĻ��ڵ�����ĸ�������
We resequenced all 22 exons, 5��UTR, 3��UTR and the exon-intron boundaries ofMCTP2 by the tagged-amplicon deep sequencing and did a single gene-based burden test for overall, female and male cohorts using the unified optimal sequence kernel association test��SKAT-O����Lee et al. 2012��. A total of 54 rare coding, four splice variants and 19 synonymous variants in NM_018349.4 with MAF �� 0.005 in EAS of 1000G, ExAC, and gnomAD were identified and verified by Sanger sequencing��Table S5��. Primary analysestested 1��all variants��disruptive variants��nonsense, essential splice site and frameshifts��plus all missense variants��154 individuals carrying��; 2��more likely to be harmful��disruptive variantsplus missense variants predicted to be harmful by at least five algorithms��79 individuals carrying��; and 3��synonymous variants��35 individuals, an individual with both a synonymous mutation and a mutation from 1��considered as a 1��carrier ��.
������֤
Mutations were amplified by PCR and validated by direct Sanger DNA sequencing. All reactions were 100% successfully validated. Primer sequences for PCR amplification are included in Table S6.
Founder Origin Testing
We genotyped nine multi-allelic microsatellite markers��CHLC.ATA22D04, AFMB077YD5, GATA128A02, AFM072YB11, AFM357TD9/D15S1038, CHLC.GATA73F01, GATA161C02, AFM217ZG1, AFM309VG9��around the c.239delG��p.S80fs��mutation ofMCTP2 from 94255659 bp to 96210769 bp on chromosome 15 in families B0001, C2149, C2666, C3030, C3049 and C3164. The inferred haplotypes were estimated by comparison to these microsatellites data collected on mother-father-offspring trios to test the possibility of the same founder origin.
����
�̼�.
In the localizer experiment, images of faces, non-face objects��e.g., chairs, food, and tools��, and texture patterns��scrambled faces��were presented at the center of the screen, subtended 6.2���6.2��. In the adaptation paradigm, the stimuli were gray-scale images of young Chinese men��hair cropped with neutral expressions��. The stimuli were presented by a MRI compatible projector system��SA-9900, The Shenzhen Sinorad Medical Electronics Co., Ltd, China, http://www.sinorad.com��, with a spatial resolution of 1,024��768 and a refresh rate of 60 Hz.
ʵ����ƺͳ���.
Each participant completed the same single scan session consisting of one functional localizer run and five runs for the adaptation paradigm. During the experiment, participants lay on their back in the scanner, using earplugs to reduce noise and sponges to hold their heads in place to reduce head movement. Participants viewed the stimuli presented on a translucent screen visible via a mirror mounted to the head coil at a distance of 60 cm. In the localizer run which lasted 360 seconds��s��, with a 12 s dummy at the beginning of the run, images appeared at a rate of 2 Hz in blocks of 12 s, interleaved with 12 s blank blocks. There were five blocks for each type of images in the run. Each image was presented for 300 milliseconds��ms��, followed by a 200 ms blank interval. Subjects performed a one-back task during scanning to ensure maintenance of attention to the stimuli. In the adaptation paradigm, pairs of face or house stimuli were presented sequentially either in the upright or the inverted manner in a randomized order that was optimized for the extraction of the hemodynamic response in an event-related fast presentation design. Each trial lasted 2000 ms. The first and second stimuli were presented for 250 ms, with an interval of 500 ms and followed by 1 s fixation. Blank screen with white cross fixation point was set between trials, with a presentation time of a random even number in the range of 0-10 s to optimize the efficiency of the event-related fMRI design by optseq2��http://surfer.nmr.mgh.harvard.edu/optseq��. Each run lasted 332 s, containing eight conditions��upright same face, upright different face, inverted same face, inverted different face, upright same house, upright different house, inverted same house and inverted different house��. Five runs were included, each containing 12 trials for each condition. Subjects made a same/different response on each trial. Here we focus only on the face stimuli.
fMRIɨ��
The fMRI data were collected in a 3T GE MR 750 scanner, with an 8-channel phase-array head coil��GE Healthcare, Waukesha, WI��at Peking University Center for MRI Research.
The gradient-echo echo planar imaging��EPI��sequence was employed for the blood oxygenation level-dependent��BOLD��signal images acquisition, and the imaging parameters were as below��repetition time��TR��= 2000 ms, echo time��TE��= 30 ms, field of view��FOV��= 224mm �� 224mm, matrix = 64 �� 64, flip angle = 90��, slice thickness = 3.5 mm with 0.7 mm spacing, voxel size = 3.5 �� 3.5 ����3.5 + 0.7��mm, 33 oblique slices covering the whole brain��.
The structure images were acquired by a 3D inversion recovery-prepped T1-weighted sequence��fSPGR, sagittal acquisition, TR = 6.65 ms, TE = 2.92 ms, TI = 450 ms, flip angle = 12��, FOV = 256 mm �� 256 mm��matrix = 256 �� 256, 192 continue slices with 1mm slice thickness, voxel size = 1 mm �� 1 mm �� 1 mm��.
��������Ϸ���
Preprocessing and data analysis were performed with SPM12��Wellcome Trust Centre for Neuroimaging, London; http://www.fil.ion.ucl.ac.uk/spm/software/spm12/��. Functional images were sequentially processedin accordance with the standard SPM approach as follows��interpolated to correct for slice timing, realigned to the middle volume, co-registered to structural scans using the mean functional image, spatially normalized to a standard echo-planar image��EPI��template based on the Montreal Neurological Institute reference brain template��MNI152, Asia brain��, and spatially smoothed with an isotropic 8 mm full width at half-maximum Gaussian kernel. For anatomical reference, the statistical maps computed were overlaid to the 3D T1-weighted scans. First, face-selective regions were localized in each subject by BOLD signals. By comparing the ��faces�� condition with the ��non-face�� condition in the localizer experiment at the first-level analysis, we assessed the face-selective region of interest��ROI��for every participant by building a 5-mm radius sphere surrounding the coordinate of the maximum activation in rFFA at a threshold p < 0.05, the family-wise error��FWE��correcting for multiple testing, with MARSBAR followed by visual confirmation of their anatomical location. For participants who lacked a face-selective area with this criterion, we explored liberal uncorrected significance thresholds��as liberal as p < 0.05��, to avoid missing effects that might be apparent at less stringent uncorrected thresholds. Because faces are processed more dominantly in the right hemisphere and because the ROIs in this study were localized more consistently in the right hemisphere, we chose to restrict our ROI-based analyses to regions in the right hemisphere��unless noted otherwise��. Second, the above-defined ROI were tested for the adaptation to facial identity using the contrast��up different faces > up identical faces��by BOLD signals at the first-level analysis. Third, the time course of percent signal change from baseline fixation was extracted from the ROI for each condition in each individual with MARSBAR, and was plotted for each condition in controls��+SD��and family members by an in-house Matlab program. Fourth, in order to directly compare each family member with the control subjects, the percent signal change in the ROI was computed for each condition. Three data points around the peak of the hemodynamic response defined individually were averaged to estimate the percent signal change. Fifth, the percent signal change was used to compute the adaptation scores��upright different face �C upright identical face��for each subject, allowing a comparison between each family member with the control group by means of Z-scores and the modified t-test score Crawford and Howell, 1998��Crawford and Garthwaite, 2002; Crawford et al., 2010��.
Results were visualized using xjView toolbox��http://www.alivelearn.net/xjview��in addition to built-in visualization in SPM12.
������ͳ�Ʒ���
Where applicable, statistical parameters including sample size, precision measures��standard error or SD��and statistical significance are reported in the figures and corresponding legends.P-values of less than 0.05 are significant.
��л
���Ǹ�л������о��������ˣ��ر�����֤�����ͥA��, ��л����ʦ����ѧ�����β�ʿ�����Ƿ���������ʶ����Ե����İ桢�Լ���������ۣ���лDarmouthѧԺ��Bradley Duchaine��ʿ����������������Ժ���������ۣ���л������ѧ��Jurg Ott�������Ժ�Taoyu Wu��ʿ���������ۣ���л������ѧ��Pianpian Zhao��ʿ�� ����ҽר��Jing Jiang��Kunjun Mao ��ʿ, �Ϸ�ҽ�ƴ�ѧ��Liang Chen��Jieyu Chen��ʿ�����ռ�����, ��л���ӱ���ʿ����ѧ��ʿ�Ա��ĵ����������ۣ���л�����廪������ѧ�������ĸ����ܼ���ƽ̨�ṩ����֧�֣���л��ҽѧ��ѧ�������ġ������Կ�ѧ�о����������廪������ѧ�������ĺ�������ʵ���ҵ�֧�֡�
�����
Abecasis, G.R., Cherny, S.S., Cookson, W.O., and Cardon, L.R.��2002��. Merlin[mdash]rapid analysis of dense genetic maps using sparse gene flow trees. Nat Genet 30, 97-101.
Adzhubei, I.A., Schmidt, S., Peshkin, L., Ramensky, V.E., Gerasimova, A., Bork, P., Kondrashov, A.S., and Sunyaev, S.R.��2010��. A method and server for predicting damaging missense mutations. Nat Methods 7, 248-249.
Afraz, S.R., Kiani, R., and Esteky, H.��2006��. Microstimulation of inferotemporal cortex influences face categorization. Nature 442, 692-695.
Allison, T., Puce, A., Spencer, D.D., and McCarthy, G.��1999��. Electrophysiological studies of human face perception. I��Potentials generated in occipitotemporal cortex by face and non-face stimuli. Cereb Cortex 9, 415-430.
Anstey, K.J., Dear, K., Christensen, H., and Jorm, A.F.��2005��. Biomarkers, health, lifestyle, and demographic variables as correlates of reaction time performance in early, middle, and late adulthood. Q J Exp Psychol A 58, 5-21.
Barraclough, N.E., and Perrett, D.I.��2011��. From single cells to social perception. Philosophical Transactions of the Royal Society B-Biological Sciences 366, 1739-1752.
Bate, S., Adams, A., Bennetts, R., and Line, H.��2019��. Developmental prosopagnosia with concurrent topographical difficulties��A case report and virtual reality training programme. Neuropsychol Rehabil 29, 1290-1312.
Bate, S., Cook, S.J., Duchaine, B., Tree, J.J., Burns, E.J., and Hodgson, T.L.��2014��. Intranasal inhalation of oxytocin improves face processing in developmental prosopagnosia. Cortex 50, 55-63.
Baylis, G.C., Rolls, E.T., and Leonard, C.M.��1985��. Selectivity between Faces in the Responses of a Population of Neurons in the Cortex in the Superior Temporal Sulcus of the Monkey. Brain Res 342, 91-102.
Behrmann, M., and Avidan, G.��2005��. Congenital prosopagnosia��face-blind from birth. Trends Cogn Sci 9, 180-187.
Bentin, S., Degutis, J.M., D'Esposito, M., and Robertson, L.C.��2007��. Too many trees to see the forest��performance, event-related potential, and functional magnetic resonance imaging manifestations of integrative congenital prosopagnosia. J Cogn Neurosci 19, 132-146.
Bodamer, J.��1947��. Die Prosop-Agnosie. Archiv f��r Psychiatrie und Nervenkrankheiten. 179, 6-53.
Bolger, A.M., Lohse, M., and Usadel, B.��2014��. Trimmomatic��a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114-2120.
Bowles, D.C., McKone, E., Dawel, A., Duchaine, B., Palermo, R., Schmalzl, L., Rivolta, D., Wilson, C.E., and Yovel, G.��2009��. Diagnosing prosopagnosia��effects of ageing, sex, and participant-stimulus ethnic match on the Cambridge Face Memory Test and Cambridge Face Perception Test. Cogn Neuropsychol 26, 423-455.
Bruce, C., Desimone, R., and Gross, C.G.��1981��. Visual Properties of Neurons in a Polysensory Area in Superior Temporal Sulcus of the Macaque. Journal of Neurophysiology 46, 369-384.
Bruce, V., and Young, A.��1986��. Understanding face recognition. Br J Psychol 77�� Pt 3��, 305-327.
Burianova, H., Lee, Y., Grady, C.L., and Moscovitch, M.��2013��. Age-related dedifferentiation and compensatory changes in the functional network underlying face processing. Neurobiol Agin g34, 2759-2767.
Busigny, T., and Rossion, B.��2010��. Acquired prosopagnosia abolishes the face inversion effect. Cortex 46, 965-981.
Carbon, C.C., Gruter, T., Gruter, M., Weber, J.E., and Lueschow, A.��2010��. Dissociation of facial attractiveness and distinctiveness processing in congenital prosopagnosia. Vis Cogn 18, 641-654.
Carbon, C.C., Gruter, T., Weber, J.E., and Lueschow, A.��2007��. Faces as objects of non-expertise��processing of thatcherised faces in congenital prosopagnosia. Perception 36, 1635-1645.
Chen, B.Q., Zhu, Z.J., Na, R., Fang, W., Zhang, W.X., Zhou, Q., Zhou, S.B., Lei, H., Huang, A.L., Chen, T.M., et al.��2018a��. Genomic Analyses of Visual Cognition��Perceptual Rivalry and Top-Down Control. Journal of Neuroscience 38, 9668-9678.
Chen, B.Q., Zhu, Z.J., Wang, Y.Y., Ding, X.H., Guo, X.B., He, M.G., Fang, W., Zhou, Q., Zhou, S.B., Lei, H., et al.��2018b��. Nature vs. nurture in human sociality��multi-level genomic analyses of social conformity. Journal of Human Genetics 63, 605-619.
Chiang, C., Layer, R.M., Faust, G.G., Lindberg, M.R., Rose, D.B., Garrison, E.P., Marth, G.T., Quinlan, A.R., and Hall, I.M.��2015��. SpeedSeq��ultra-fast personal genome analysis and interpretation. Nat Methods 12, 966-968.
Chitty, A.J., Perrett, D.I., Mistlin, A.J., and Potter, D.D.��1985��. Visual Cells in the Temporal Cortex Selectively Responsive to the Sight of Hands Manipulating Objects. Perception 14, A15-A15.
Cho, W., and Stahelin, R.V.��2006��. Membrane binding and subcellular targeting of C2 domains. Biochimica et Biophysica Acta��BBA��- Molecular and Cell Biology of Lipids1761, 838-849.
Cirulli, E.T., and Goldstein, D.B.��2010��. Uncovering the roles of rare variants in common disease through whole-genome sequencing. Nat Rev Genet 11, 415-425.
Clark, V.P., Keil, K., Maisog, J.M., Courtney, S., Ungerleider, L.G., and Haxby, J.V.��1996��. Functional magnetic resonance imaging of human visual cortex during face matching��A comparison with positron emission tomography. Neuroimage 4, 1-15.
Cong, P.K., Bai, W.Y., Li, J.C., Yang, M.Y., Khederzadeh, S., Gai, S.R., Li, N., Liu, Y.H., Yu, S.H., Zhao, W.W., et al.��2022a��. Genomic analyses of 10,376 individuals in the Westlake BioBank for Chinese��WBBC��pilot project. Nat Commun 13, 2939.
Cong, P.K., Khederzadeh, S., Yuan, C.D., Ma, R.J., Zhang, Y.Y., Liu, J.Q., Yu, S.H., Xu, L., Gao, J.H., Pan, H.X., et al.��2022b��. Identification of clinically actionable secondary genetic variants from whole-genome sequencing in a large-scale Chinese population. Clin Transl Med 12, e866.
Crawford, J.R., and Garthwaite, P.H.��2002��. Investigation of the single case in neuropsychology��confidence limits on the abnormality of test scores and test score differences. Neuropsychologia 40, 1196-1208.
Crawford, J.R., Garthwaite, P.H., and Porter, S.��2010��. Point and interval estimates of effect sizes for the case-controls design in neuropsychology��rationale, methods, implementations, and proposed reporting standards. Cogn Neuropsychol 27, 245-260.
Crawford, J.R., and Howell, D.C.��1998��. Comparing an individual's test score against norms derived from small samples. Clinical Neuropsychologist 12, 482-486.
Dalrymple, K.A., and Palermo, R.��2016��. Guidelines for studying developmental prosopagnosia in adults and children. Wiley Interdiscip Rev Cogn Sci 7, 73-87.
Damasio, A.R., Tranel, D., and Damasio, H.��1990��. Face agnosia and the neural substrates of memory. Annu Rev Neurosci 13, 89-109.
De Haan, E.H.��1999��. A familial factor in the development of face recognition deficits. J Clin Exp Neuropsychol21, 312-315.
Dennett, H.W., McKone, E., Tavashmi, R., Hall, A., Pidcock, M., Edwards, M., and Duchaine, B.��2012��. The Cambridge Car Memory Test��a task matched in format to the Cambridge Face Memory Test, with norms, reliability, sex differences, dissociations from face memory, and expertise effects. Behav Res Methods 44, 587-605.
Desimone, R., Albright, T.D., Gross, C.G., and Bruce, C.��1984��. Stimulus-Selective Properties of Inferior Temporal Neurons in the Macaque. Journal of Neuroscience 4, 2051-2062.
Dinkelacker, V., Gruter, M., Klaver, P., Gruter, T., Specht, K., Weis, S., Kennerknecht, I., Elger, C.E., and Fernandez, G.��2011��. Congenital prosopagnosia��multistage anatomical and functional deficits in face processing circuitry. J Neurol 258, 770-782.
Djurovic, S., Le Hellard, S., Kahler, A.K., Jonsson, E.G., Agartz, I., Steen, V.M., Hall, H., Wang, A.G., Rasmussen, H.B., Melle, I., et al.��2009��. Association of MCTP2 gene variants with schizophrenia in three independent samples of Scandinavian origin��SCOPE��. Psychiatry Res 168, 256-258.
Dobel, C., Bolte, J., Aicher, M., and Schweinberger, S.R.��2007��. Prosopagnosia without apparent cause��overview and diagnosis of six cases. Cortex 43, 718-733.
Dricot, L., Sorger, B., Schiltz, C., Goebel, R., and Rossion, B.��2008��. The roles of ��face�� and ��non-face�� areas during individual face perception��Evidence by fMRI adaptation in a brain-damaged prosopagnosic patient. NeuroImage 40, 318-332.
Duchaine, B., Germine, L., and Nakayama, K.��2007��. Family resemblance��ten family members with prosopagnosia and within-class object agnosia. Cogn Neuropsychol 24, 419-430.
Duchaine, B., and Nakayama, K.��2006a��. The Cambridge Face Memory Test��results for neurologically intact individuals and an investigation of its validity using inverted face stimuli and prosopagnosic participants. Neuropsychologia 44, 576-585.
Duchaine, B.C., and Nakayama, K.��2004��. Developmental prosopagnosia and the Benton Facial Recognition Test. Neurology 62, 1219-1220.
Duchaine, B.C., and Nakayama, K.��2006b��. Developmental prosopagnosia��a window to content-specific face processing. Curr Opin Neurobiol 16, 166-173.
Duchaine, B.C., and Weidenfeld, A.��2003��. An evaluation of two commonly used tests of unfamiliar face recognition. Neuropsychologia 41, 713-720.
Dunning, D., Johnson, K., Ehrlinger, J., and Kruger, J.��2003��. Why People Fail to Recognize Their Own Incompetence. 12, 83-87.
Dzhelyova, M.P., Ellison, A., and Atkinson, A.P.��2011��. Event-related repetitive TMS reveals distinct, critical roles for right OFA and bilateral posterior STS in judging the sex and trustworthiness of faces. J Cogn Neurosci 23, 2782-2796.
Esins, J., Schultz, J., Stemper, C., Kennerknecht, I., and Bulthoff, I.��2016��. Face Perception and Test Reliabilities in Congenital Prosopagnosia in Seven Tests. Iperception 7, 2041669515625797.
Esins, J., Schultz, J., Stemper, C., Kennerknecht, I., Wallraven, C., and Bulthoff, I.��2015��. Corrigendum��Do congenital prosopagnosia and the other-race effect affect the same face recognition mechanisms? Front Hum Neurosci 9, 294.
Espino-Saldaña, A.E., Dur��n-R��os, K., Olivares-Hernandez, E., Rodr��guez-Ortiz, R., Arellano-Carbajal, F., and Mart��nez-Torres, A.��2020��. Temporal and spatial expression of zebrafish mctp genes and evaluation of frameshift alleles of mctp2b. Gene 738, 144371.
Forshew, T., Murtaza, M., Parkinson, C., Gale, D., Tsui, D.W., Kaper, F., Dawson, S.J., Piskorz, A.M., Jimenez-Linan, M., Bentley, D., et al.��2012��. Noninvasive identification and monitoring of cancer mutations by targeted deep sequencing of plasma DNA. Sci Transl Med 4, 136ra168.
Freiwald, W., Duchaine, B., and Yovel, G.��2016��. Face Processing Systems��From Neurons to Real-World Social Perception. Annu Rev Neurosci 39, 325-346.
Galaburda, A.M., and Duchaine, B.C.��2003��. Developmental disorders of vision. Neurol Clin 21, 687-707.
Gao, X., Vuong, Q.C., and Rossion, B.��2019��. The cortical face network of the prosopagnosic patient PS with fast periodic stimulation in fMRI. Cortex 119, 528-542.
Genc, O., Dickman, D.K., Ma, W., Tong, A., Fetter, R.D., and Davis, G.W.��2017��. MCTP is an ER-resident calcium sensor that stabilizes synaptic transmission and homeostatic plasticity. Elife6.
Genomes Project, C., Abecasis, G.R., Altshuler, D., Auton, A., Brooks, L.D., Durbin, R.M., Gibbs, R.A., Hurles, M.E., and McVean, G.A.��2010��. A map of human genome variation from population-scale sequencing. Nature4 67, 1061-1073.
Genomes Project, C., Abecasis, G.R., Auton, A., Brooks, L.D., DePristo, M.A., Durbin, R.M., Handsaker, R.E., Kang, H.M., Marth, G.T., and McVean, G.A.��2012��. An integrated map of genetic variation from 1,092 human genomes. Nature 491, 56-65.
Genomes Project, C., Auton, A., Brooks, L.D., Durbin, R.M., Garrison, E.P., Kang, H.M., Korbel, J.O., Marchini, J.L., McCarthy, S., McVean, G.A., et al.��2015��. A global reference for human genetic variation. Nature 526, 68-74.
Germine, L.T., Duchaine, B., and Nakayama, K.��2011��. Where cognitive development and aging meet��face learning ability peaks after age 30. Cognition 118, 201-210.
Goh, J.O., Suzuki, A., and Park, D.C.��2010��. Reduced neural selectivity increases fMRI adaptation with age during face discrimination. Neuroimage 51, 336-344.
Grill-Spector, K., Henson, R., and Martin, A.��2006��. Repetition and the brain��neural models of stimulus-specific effects. Trends Cogn Sci 10, 14-23.
Grill-Spector, K., Knouf, N., and Kanwisher, N.��2004��. The fusiform face area subserves face perception, not generic within-category identification. Nat Neurosci 7, 555-562.
Grill-Spector, K., and Malach, R.��2001��. fMR-adaptation��a tool for studying the functional properties of human cortical neurons. Acta Psychol��Amst��107, 293-321.
Grill-Spector, K., and Malach, R.��2004��. The human visual cortex. Annu Rev Neurosci 27, 649-677.
Gross, C.G., Rochamir.Ce, and Bender, D.B.��1972��. Visual Properties of Neurons in Inferotemporal Cortex of Macaque. Journal of Neurophysiology 35, 96-&.
Grueter, M., Grueter, T., Bell, V., Horst, J., Laskowski, W., Sperling, K., Halligan, P.W., Ellis, H.D., and Kennerknecht, I.��2007��. Hereditary prosopagnosia��the first case series. Cortex 43, 734-749.
Gruter, T., Gruter, M., Bell, V., and Carbon, C.C.��2009��. Visual mental imagery in congenital prosopagnosia. Neurosci Lett 453, 135-140.
Gruter, T., Gruter, M., and Carbon, C.C.��2008��. Neural and genetic foundations of face recognition and prosopagnosia. J Neuropsychol 2, 79-97.
Gusella, J.F., Wexler, N.S., Conneally, P.M., Naylor, S.L., Anderson, M.A., Tanzi, R.E., Watkins, P.C., Ottina, K., Wallace, M.R., Sakaguchi, A.Y., et al.��1983��. A polymorphic DNA marker genetically linked to Huntington's disease. Nature 306, 234-238.
Hadjikhani, N., and de Gelder, B.��2002��. Neural basis of prosopagnosia��an fMRI study. Hum Brain Mapp 16, 176-182.
Haxby, J.V., Hoffman, E.A., and Gobbini, M.I.��2000��. The distributed human neural system for face perception. Trends Cogn Sci 4, 223-233.
Haxby, J.V., Horwitz, B., Ungerleider, L.G., Maisog, J.M., Pietrini, P., and Grady, C.L.��1994��. The Functional-Organization of Human Extrastriate Cortex - a Pet-Rcbf Study of Selective Attention to Faces and Locations. Journal of Neuroscience 14, 6336-6353.
Heinzen, E.L., Neale, B.M., Traynelis, S.F., Allen, A.S., and Goldstein, D.B.��2015��. The genetics of neuropsychiatric diseases��looking in and beyond the exome. Annu Rev Neurosci 38, 47-68.
Henson, R.N., and Rugg, M.D.��2003��. Neural response suppression, haemodynamic repetition effects, and behavioural priming. Neuropsychologia 41, 263-270.
Hung, C.C., Yen, C.C., Ciuchta, J.L., Papoti, D., Bock, N.A., Leopold, D.A., and Silva, A.C.��2015��. Functional mapping of face-selective regions in the extrastriate visual cortex of the marmoset. J Neurosci 35, 1160-1172.
Johnen, A., Schmukle, S.C., Huttenbrink, J., Kischka, C., Kennerknecht, I., and Dobel, C.��2014��. A family at risk��congenital prosopagnosia, poor face recognition and visuoperceptual deficits within one family. Neuropsychologia 58, 52-63.
Jordan, D.M., Ramensky, V.E., and Sunyaev, S.R.��2010��. Human allelic variation��perspective from protein function, structure, and evolution. Curr Opin Struct Biol 20, 342-350.
Kanwisher, N., McDermott, J., and Chun, M.M.��1997��. The fusiform face area��a module in human extrastriate cortex specialized for face perception. J Neurosci 17, 4302-4311.
Kennerknecht, I.��2021��. Prevalence of Hereditary Prosopagnosia �C A Worldwide Survey. . M��nster��Westfälische Wilhelms-Universität M��nster
Kennerknecht, I., Grueter, T., Welling, B., Wentzek, S., Horst, J., Edwards, S., and Grueter, M.��2006��. First report of prevalence of non-syndromic hereditary prosopagnosia��HPA��. Am J Med Genet A 140, 1617-1622.
Kennerknecht, I., Ho, N.Y., and Wong, V.C.��2008a��. Prevalence of hereditary prosopagnosia��HPA��in Hong Kong Chinese population. Am J Med Genet A 146A, 2863-2870.
Kennerknecht, I., Pluempe, N., and Welling, B.��2008b��. Congenital prosopagnosia--a common hereditary cognitive dysfunction in humans. Front Biosci 13, 3150-3158.
Kennerknecht, I., Plumpe, N., Edwards, S., and Raman, R.��2007��. Hereditary prosopagnosia��HPA����the first report outside the Caucasian population. Journal of Human Genetics 52, 230-236.
Kreiman, G., Koch, C., and Fried, I.��2000��. Category-specific visual responses of single neurons in the human medial temporal lobe. Nat Neurosci 3, 946-953.
Kress, T., and Daum, I.��2003��. Developmental prosopagnosia��a review. Behav Neurol14, 109-121.
Kruger, J., and Dunning, D.��1999��. Unskilled and unaware of it��how difficulties in recognizing one's own incompetence lead to inflated self-assessments. J Pers Soc Psychol 77, 1121-1134.
Kweon, K., Shin, E.S., Park, K.J., Lee, J.K., Joo, Y., and Kim, H.W.��2018��. Genome-Wide Analysis Reveals Four Novel Loci for Attention-Deficit Hyperactivity Disorder in Korean Youths. Soa--ch'ongsonyon chongsin uihak = Journal of child & adolescent psychiatry 29, 62-72.
Le Grand, R., Cooper, P.A., Mondloch, C.J., Lewis, T.L., Sagiv, N., de Gelder, B., and Maurer, D.��2006��. What aspects of face processing are impaired in developmental prosopagnosia? Brain Cogn 61, 139-158.
Lee, S., Wu, M.C., and Lin, X.��2012��. Optimal tests for rare variant effects in sequencing association studies. Biostatistics 13, 762-775.
Lee, Y., Duchaine, B., Wilson, H.R., and Nakayama, K.��2010��. Three cases of developmental prosopagnosia from one family��detailed neuropsychological and psychophysical investigation of face processing. Cortex 46, 949-964.
Lee, Y., Grady, C.L., Habak, C., Wilson, H.R., and Moscovitch, M.��2011��. Face processing changes in normal aging revealed by fMRI adaptation. J Cogn Neurosci 23, 3433-3447.
Lek, M., Karczewski, K.J., Minikel, E.V., Samocha, K.E., Banks, E., Fennell, T., O'Donnell-Luria, A.H., Ware, J.S., Hill, A.J., Cummings, B.B., et al.��2016��. Analysis of protein-coding genetic variation in 60,706 humans. Nature 536, 285-291.
Li, H., and Durbin, R.��2009��. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754-1760.
Liu, J., Harris, A., and Kanwisher, N.��2010��. Perception of Face Parts and Face Configurations��An fMRI Study. Journal of Cognitive Neuroscience 22, 203-211.
Malach, R., Reppas, J.B., Benson, R.R., Kwong, K.K., Jiang, H., Kennedy, W.A., Ledden, P.J., Brady, T.J., Rosen, B.R., and Tootell, R.B.H.��1995��. Object-Related Activity Revealed by Functional Magnetic-Resonance-Imaging in Human Occipital Cortex. P Natl Acad Sci USA 92, 8135-8139.
Manichaikul, A., Mychaleckyj, J.C., Rich, S.S., Daly, K., Sale, M., and Chen, W.M.��2010��. Robust relationship inference in genome-wide association studies. Bioinformatics 26, 2867-2873.
McCall, M.N., Uppal, K., Jaffee, H.A., Zilliox, M.J., and Irizarry, R.A.��2011��. The Gene Expression Barcode��leveraging public data repositories to begin cataloging the human and murine transcriptomes. Nucleic Acids Res 39, D1011-1015.
McCarthy, G., Puce, A., Belger, A., and Allison, T.��1999��. Electrophysiological studies of human face perception. II��Response properties of face-specific potentials generated in occipitotemporal cortex. Cereb Cortex 9, 431-444.
McCarthy, G., Puce, A., Gore, J.C., and Allison, T.��1997��. Face-specific processing in the human fusiform gyrus. J Cogn Neurosci 9, 605-610.
McConachie, H.R.��1976��. Developmental prosopagnosia. A single case report. Cortex 12, 76-82.
McKenna, A., Hanna, M., Banks, E., Sivachenko, A., Cibulskis, K., Kernytsky, A., Garimella, K., Altshuler, D., Gabriel, S., Daly, M., et al.��2010��. The Genome Analysis Toolkit��a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res 20, 1297-1303.
McKone, E., and Palermo, R.��2010��. A strong role for nature in face recognition. Proc Natl Acad Sci U S A 107, 4795-4796.
McKone, E., Stokes, S., Liu, J., Cohan, S., Fiorentini, C., Pidcock, M., Yovel, G., Broughton, M., and Pelleg, M.��2012��. A robust method of measuring other-race and other-ethnicity effects��the Cambridge Face Memory Test format. PLoS One 7, e47956.
Mick, E., Todorov, A., Smalley, S., Hu, X., Loo, S., Todd, R.D., Biederman, J., Byrne, D., Dechairo, B., Guiney, A., et al.��2010��. Family-Based Genome-Wide Association Scan of Attention-Deficit/Hyperactivity Disorder. Journal of the American Academy of Child & Adolescent Psychiatry 49, 898-905.e893.
Moeller, S., Crapse, T., Chang, L., and Tsao, D.Y.��2017��. The effect of face patch microstimulation on perception of faces and objects. Nat Neurosci 20, 743-+.
Moeller, S., Freiwald, W.A., and Tsao, D.Y.��2008��. Patches with links��a unified system for processing faces in the macaque temporal lobe. Science 320, 1355-1359.
Ng, P.C., and Henikoff, S.��2001��. Predicting deleterious amino acid substitutions. Genome Res 11, 863-874.
Nunn, J.A., Postma, P., and Pearson, R.��2001��. Developmental prosopagnosia��should it be taken at face value? Neurocase 7, 15-27.
Park, J., Carp, J., Kennedy, K.M., Rodrigue, K.M., Bischof, G.N., Huang, C.M., Rieck, J.R., Polk, T.A., and Park, D.C.��2012��. Neural broadening or neural attenuation? Investigating age-related dedifferentiation in the face network in a large lifespan sample. J Neurosci 32, 2154-2158.
Perrett, D.I., Mistlin, A.J., and Chitty, A.J.��1987��. Visual Neurons Responsive to Faces. Trends in Neurosciences 10, 358-364.
Perrett, D.I., Rolls, E.T., and Caan, W.��1982��. Visual Neurones Responsive to Faces in the Monkey Temporal Cortex. Exp Brain Res 47, 329-342.
Perrett, D.I., Smith, P.A.J., Potter, D.D., Mistlin, A.J., Head, A.S., Milner, A.D., and Jeeves, M.A.��1984��. Neurons Responsive to Faces in the Temporal Cortex - Studies of Functional-Organization, Sensitivity to Identity and Relation to Perception. Human Neurobiology 3, 197-208.
Perrett, D.I., Smith, P.A.J., Potter, D.D., Mistlin, A.J., Head, A.S., Milner, A.D., and Jeeves, M.A.��1985��. Visual Cells in the Temporal Cortex Sensitive to Face View and Gaze Direction. Proceedings of the Royal Society Series B-Biological Sciences 223, 293-317.
Pitcher, D., Charles, L., Devlin, J.T., Walsh, V., and Duchaine, B.��2009��. Triple Dissociation of Faces, Bodies, and Objects in Extrastriate Cortex. Curr Biol 19, 319-324.
Pitcher, D., Garrido, L., Walsh, V., and Duchaine, B.C.��2008��. Transcranial magnetic stimulation disrupts the perception and embodiment of facial expressions. J Neurosci 28, 8929-8933.
Poelmans, G., Pauls, D.L., Buitelaar, J.K., and Franke, B.��2011��. Integrated genome-wide association study findings��identification of a neurodevelopmental network for attention deficit hyperactivity disorder. Am J Psychiat 168, 365-377.
Polk, T.A., Park, J., Smith, M.R., and Park, D.C.��2007��. Nature versus nurture in ventral visual cortex��A functional magnetic resonance Imaging study of twins. J Neurosci 27, 13921-13925.
Pritchard, J.K.��2001��. Are rare variants responsible for susceptibility to complex diseases? Am J Hum Genet 69, 124-137.
Puce, A., Allison, T., Asgari, M., Gore, J.C., and McCarthy, G.��1996��. Differential Sensitivity of Human Visual Cortex to Faces, Letterstrings, and Textures��A Functional Magnetic Resonance Imaging Study. J Neurosci 16, 5205-5215.
Puce, A., Allison, T., and McCarthy, G.��1999��. Electrophysiological studies of human face perception. III��Effects of top-down processing on face-specific potentials. Cereb Cortex 9, 445-458.
Purcell, S., Neale, B., Todd-Brown, K., Thomas, L., Ferreira, M.A., Bender, D., Maller, J., Sklar, P., de Bakker, P.I., Daly, M.J., et al.��2007��. PLINK��a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet 81, 559-575.
Purcell, S.M., Moran, J.L., Fromer, M., Ruderfer, D., Solovieff, N., Roussos, P., O'Dushlaine, C., Chambert, K., Bergen, S.E., Kahler, A., et al.��2014��. A polygenic burden of rare disruptive mutations in schizophrenia. Nature 506, 185-190.
Qiu, L., Yu, H., and Liang, F.��2015��. Multiple C2 domains transmembrane protein 1 is expressed in CNS neurons and possibly regulates cellular vesicle retrieval and oxidative stress. J Neurochem 135, 492-507.
Rangarajan, V., Hermes, D., Foster, B.L., Weiner, K.S., Jacques, C., Grill-Spector, K., and Parvizi, J.��2014��. Electrical stimulation of the left and right human fusiform gyrus causes different effects in conscious face perception. J Neurosci 34, 12828-12836.
Rasetti, R., and Weinberger, D.R.��2011��. Intermediate phenotypes in psychiatric disorders. Curr Opin Genet Dev 21, 340-348.
Righart, R., Andersson, F., Schwartz, S., Mayer, E., and Vuilleumier, P.��2009��. Top-Down Activation of Fusiform Cortex without Seeing Faces in Prosopagnosia. Cerebral Cortex 20, 1878-1890.
Robinson, J.T., Thorvaldsdottir, H., Winckler, W., Guttman, M., Lander, E.S., Getz, G., and Mesirov, J.P.��2011��. Integrative genomics viewer. Nat Biotechnol 29, 24-26.
Rolls, E.T.��1984��. Neurons in the Cortex of the Temporal-Lobe and in the Amygdala of the Monkey with Responses Selective for Faces. Hum Neurobiol 3, 209-222.
Rolls, E.T., Baylis, G.C., and Leonard, C.M.��1985��. Role of Low and High Spatial-Frequencies in the Face-Selective Responses of Neurons in the Cortex in the Superior Temporal Sulcus in the Monkey. Vision Res 25, 1021-1035.
Rossion, B., Dricot, L., Goebel, R., and Busigny, T.��2011��. Holistic Face Categorization in Higher Order Visual Areas of the Normal and Prosopagnosic Brain��Toward a Non-Hierarchical View of Face Perception. 4.
Rotshtein, P., Henson, R.N., Treves, A., Driver, J., and Dolan, R.J.��2005��. Morphing Marilyn into Maggie dissociates physical and identity face representations in the brain. Nat Neurosci 8, 107-113.
Saito, H., Yukie, M., Tanaka, K., Hikosaka, K., Fukada, Y., and Iwai, E.��1986��. Integration of Direction Signals of Image Motion in the Superior Temporal Sulcus of the Macaque Monkey. Journal of Neuroscience 6, 145-157.
Schiltz, C., and Rossion, B.��2006��. Faces are represented holistically in the human occipito-temporal cortex. Neuroimage 32, 1385-1394.
Schiltz, C., Sorger, B., Caldara, R., Ahmed, F., Mayer, E., Goebel, R., and Rossion, B.��2006��. Impaired face discrimination in acquired prosopagnosia is associated with abnormal response to individual faces in the right middle fusiform gyrus. Cereb Cortex 16, 574-586.
Schmalzl, L., Palermo, R., and Coltheart, M.��2008��. Cognitive heterogeneity in genetically based prosopagnosia��a family study. J Neuropsycho l2, 99-117.
Schrott, H.G., Goldstein, J.L., Hazzard, W.R., McGoodwin, M.M., and Motulsky, A.G.��1972��. Familial hypercholesterolemia in a large indred. Evidence for a monogenic mechanism. Ann Intern Med 76, 711-720.
Scott, L.J., Muglia, P., Kong, X.Q., Guan, W., Flickinger, M., Upmanyu, R., Tozzi, F., Li, J.Z., Burmeister, M., Absher, D., et al.��2009��. Genome-wide association and meta-analysis of bipolar disorder in individuals of European ancestry. Proc Natl Acad Sci U S A 106, 7501-7506.
Sergent, J., Ohta, S., and Macdonald, B.��1992��. Functional Neuroanatomy of Face and Object Processing - a Positron Emission Tomography Study. Brain115, 15-36.
Shah, P., Gaule, A., Sowden, S., Bird, G., and Cook, R.��2015��. The 20-item prosopagnosia index��PI20����a self-report instrument for identifying developmental prosopagnosia. R Soc Open Sci 2, 140343.
Sherry, S.T., Ward, M.H., Kholodov, M., Baker, J., Phan, L., Smigielski, E.M., and Sirotkin, K.��2001��. dbSNP��the NCBI database of genetic variation. Nucleic Acids Res 29, 308-311.
Shin, O.H., Han, W., Wang, Y., and Sudhof, T.C.��2005��. Evolutionarily conserved multiple C2 domain proteins with two transmembrane regions��MCTPs��and unusual Ca2+ binding properties. J Biol Chem 280, 1641-1651.
Shupliakov, O., and Brodin, L.��2010��. Recent insights into the building and cycling of synaptic vesicles. Exp Cell Res 316, 1344-1350.
Sjostedt, E., Zhong, W., Fagerberg, L., Karlsson, M., Mitsios, N., Adori, C., Oksvold, P., Edfors, F., Limiszewska, A., Hikmet, F., et al.��2020��. An atlas of the protein-coding genes in the human, pig, and mouse brain. Science 367.
Stollhoff, R., Jost, J., Elze, T., and Kennerknecht, I.��2010��. The early time course of compensatory face processing in congenital prosopagnosia. PLoS One 5, e11482.
Stollhoff, R., Jost, J., Elze, T., and Kennerknecht, I.��2011��. Deficits in long-term recognition memory reveal dissociated subtypes in congenital prosopagnosia. PLoS One 6, e15702.
Susilo, T., and Duchaine, B.��2013��. Advances in developmental prosopagnosia research. Current Opinion in Neurobiology 23, 423-429.
Thorvaldsdottir, H., Robinson, J.T., and Mesirov, J.P.��2013��. Integrative Genomics Viewer��IGV����high-performance genomics data visualization and exploration. Brief Bioinform 14, 178-192.
Tsao, D.Y., Freiwald, W.A., Knutsen, T.A., Mandeville, J.B., and Tootell, R.B.��2003��. Faces and objects in macaque cerebral cortex. Nat Neurosci 6, 989-995.
Tsao, D.Y., Freiwald, W.A., Tootell, R.B., and Livingstone, M.S.��2006��. A cortical region consisting entirely of face-selective cells. Science 311, 670-674.
Tsao, D.Y., and Livingstone, M.S.��2008��. Mechanisms of face perception. Annu Rev Neurosci 31, 411-437.
Tsao, D.Y., Moeller, S., and Freiwald, W.A.��2008��. Comparing face patch systems in macaques and humans. Proc Natl Acad Sci U S A 105, 19514-19519.
Tsui, L.C., Buchwald, M., Barker, D., Braman, J.C., Knowlton, R., Schumm, J.W., Eiberg, H., Mohr, J., Kennedy, D., Plavsic, N., et al.��1985��. Cystic fibrosis locus defined by a genetically linked polymorphic DNA marker. Science 230, 1054-1057.
Verfaillie, K., Huysegems, S., De Graef, P., and Van Belle, G.��2014��. Impaired holistic and analytic face processing in congenital prosopagnosia��Evidence from the eye-contingent mask/window paradigm. Vis Cogn 22, 503-521.
von Kriegstein, K., Dogan, O., Gruter, M., Giraud, A.L., Kell, C.A., Gruter, T., Kleinschmidt, A., and Kiebel, S.J.��2008��. Simulation of talking faces in the human brain improves auditory speech recognition. Proc Natl Acad Sci U S A 105, 6747-6752.
Wang, K., Li, M., Hadley, D., Liu, R., Glessner, J., Grant, S.F., Hakonarson, H., and Bucan, M.��2007��. PennCNV��an integrated hidden Markov model designed for high-resolution copy number variation detection in whole-genome SNP genotyping data. Genome Res 17, 1665-1674.
Wang, K., Li, M., and Hakonarson, H.��2010��. ANNOVAR��functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res 38, e164.
White, D., and Burton, A.M.��2022��. Individual differences and the multidimensional nature of face perception. Nature Reviews Psychology 1, 287-300.
Wilmer, J.B., Germine, L., Chabris, C.F., Chatterjee, G., Williams, M., Loken, E., Nakayama, K., and Duchaine, B.��2010��. Human face recognition ability is specific and highly heritable. P Natl Acad Sci USA 107, 5238-5241.
Yamane, S., Kaji, S., and Kawano, K.��1988��. What Facial Features Activate Face Neurons in the Inferotemporal Cortex of the Monkey. Exp Brain Res 73, 209-214.
Yovel, G., and Kanwisher, N.��2004��. Face perception��domain specific, not process specific. Neuron 44, 889-898.
Yovel, G., and Kanwisher, N.��2005��. The neural basis of the behavioral face-inversion effect. Curr Biol 15, 2256-2262.
Zebrowitz, L., Ward, N., Boshyan, J., Gutchess, A., and Hadjikhani, N.��2016��. Dedifferentiated face processing in older adults is linked to lower resting state metabolic activity in fusiform face area. Brain Res 1644, 22-31.
Zhang, J., Li, X., Song, Y., and Liu, J.��2012��. The fusiform face area is engaged in holistic, not parts-based, representation of faces. PLoS One7, e40390.
Zhao, Y., Zhen, Z., Liu, X., Song, Y., and Liu, J.��2018��. The neural network for face recognition��Insights from an fMRI study on developmental prosopagnosia. Neuroimage 169, 151-161.
Zhao, Y.F., Li, J.G., Liu, X.Q., Song, Y.Y., Wang, R.S., Yang, Z.T., and Liu, J.��2016��. Altered spontaneous neural activity in the occipital face area reflects behavioral deficits in developmental prosopagnosia. Neuropsychologia 89, 344-355.
Zhou, X., and Jenkins, R.��2020��. Dunning-Kruger effects in face perception. Cognition 203, 104345.
Zhu, Q., Song, Y.Y., Hu, S.Y., Li, X.B., Tian, M.Q., Zhen, Z.L., Dong, Q., Kanwisher, N., and Liu, J.��2010��. Heritability of the Specific Cognitive Ability of Face Perception. Current Biology 20, 137-142.
Zhu, X.W., Liu, K.Q., Wang, P.Y., Liu, J.Q., Chen, J.Y., Xu, X.J., Xu, J.J., Qiu, M.C., Sun, Y., Liu, C., et al.��2021��. Cohort profile��the Westlake BioBank for Chinese��WBBC��pilot project. BMJ Open 11, e045564.
Zhu, Z., Chen, B., Na, R., Fang, W., Zhang, W., Zhou, Q., Zhou, S., Lei, H., Huang, A., Chen, T., et al.��2019��. Heritability of human visual contour integration-an integrated genomic study. Eur J Hum Genet 27, 1867-1875.
Zhu, Z., Wang, Y., Cao, Z., Chen, B., Cai, H., Wu, Y., and Rao, Y.��2016��. Cue-independent memory impairment by reactivation-coupled interference in human declarative memory. Cognition 155, 125-134.
Zhu, Z.J., Chen, B.Q., Yan, H.M., Fang, W., Zhou, Q., Zhou, S.B., Lei, H., Huang, A.L., Chen, T.M., Gao, T.M., et al.��2018��. Multi-level genomic analyses suggest new genetic variants involved in human memory. Eur J Hum Genet 26, 1668-1678.
Zhu, Z., Chen, B.Q., Na, R., Fang, W., Zhang, W.X., Zhou, Q., Zhou, S.B., Lei, H., Huang, A.L., Chen, T.M., Gao, T.M., Chen, L., Ni, D.S., Gu., Y.P., Liu, J.N., Chen, J.Y., Rao, Y. and Fang, F.��2020��. Heritability of human visual controur integration--an integrated genomic study. Eur J Hum Genet 27, 1867-1875.
ͼ��
ͼ1��Mapping of a Novel Autosomal Dominant CP Locus to the Short Arm of Chromosome 15��15q26��.��a��The pedigree plot of family A with genotypes for c.2147T>G��p.I716S��, GenBank��NM_018349 and phenotypes. Crosses, DNA samples not available; diagonal slashes, dead members; filled symbols, family members poor on both interviews and the behavioral CFMT test ; half-filled symbols, family members who had daily face recognition problems but appeared normal on CFMT or founders with daily face recognition problems ; an arrow head, the index of family A.
��b��Graphical representation of parametric linkage results of linkage 1 on chr15��LOD score = 3.49��in family A with diagnosis based only on interviews as well as the CFMT behavioral test, assuming a rare dominant model.
��c��Graphical representation of parametric linkage results of linkage 2 on chr15��LOD score = 5.13��in family A with diagnosis based on interviews not behavioral tests, assuming a rare dominant model.
ͼ2��Mutations Detection in MCTP2 and Co-segregation in CP.
��a��The distribution of the 20-item questionnaire scores in 743 males��the blue approximate normal curve��and 2161 females��the red approximate normal curve��. Dashed lines indicate the cut-off of 2 SDs poorer than the mean. Sticks with different colors indicate different individuals with different mutations.
��b��A mutation c.239delG, leading to p.S80fs in individuals B0001, B0009, B0046 with CP. All three of them were aware of difficulties with face recognition early in their life��Table S2��.
��c��Correlation of the��c.239delG, p.S80fs��genotype with the CP phenotype in the family of B0001, with 3 members tested.
��d��A mutation c.814A>T, leading to p. M272L in individual B0011.
��e��Correlation of the genotype��c.814A>T [p.M272L]��with the CP phenotype in the family of B0011.
��f��A mutation c.1120A>G, leading to p.T374A in individual B0010.
��g��A mutation c.1642G>A, leading to p.V548I in individual B0003.
��h��Correlation of the��c.1642G>A [p.V548I]��genotype with the CP phenotype in the family of B0003, with 6 members tested��Table S2��.
��i��A mutation c.1922G>A, leading to p.R641Q in individual B0002.
��j��Correlation of the��c.1922G>A [p.R641Q]��genotype with the CP phenotype in the family of B0002, with 5 members tested��Table S2��.
ͼ3��Co-segregation of c.239delG��p.S80fs��in MCTP2 in CP.
16 individuals with the frameshift deletion mutation��c.239delG [p.S80fs]��were detected in a cohort of 1757 students. 14 of them were available for further analysis. More members from 6 families were available��Table S3��.
��a��Correlation of the��c.239delG [p.S80fs]��genotype with the CP phenotype in the family of C2666, with 10 family members tested.
��b��Correlation of the��c.239delG [p.S80fs]��genotype with the CP phenotype in the family of C3164, with 4 family members tested.
��c��Correlation of the��c.239delG [p.S80fs]��genotype with the CP phenotype in the family of C3049 with 6 family members tested.
��d��Correlation of the��c.239delG [p.S80fs]��genotype with the CP phenotype in the family of C2149 with 4 family members tested.
��e��Correlation of the��c.239delG [p.S80fs]��genotype with the CP phenotype in the family of C3030, with 3 members tested.
��f��The pedigree plot of family C3649 with genotype and phenotype.
ͼ4��rFFA Activation in Face Discrimination
Experimental paradigm as Figure 4a. The average percent signal change����SD for controls��from baseline fixation is plotted for the different and identical upright face conditions. Bold response to pairs of upright different faces or identical faces in the rFFA of controls��n=20����b��; non-CP members A-VI:1��c��, A-VI:2��d��, A-VI:10��e��and CP members A-VI:5��f��, A-VI:7��g��, and A-VI:9��h��from family A with the I716S mutation inMCTP2; non-CP C2666-III:2��i��and CP C2666-III:4��j��from family C2666 with the S80fs mutation in MCTP2.
ͼ5��Comparison between CP and Control Individuals in the Adaptation Experiment
The adaptation score��difference in percent signal change between responses to upright different faces and upright identical faces��is plotted for CP individuals��black bars��and each individual control subject��white bars��in an increasing order.
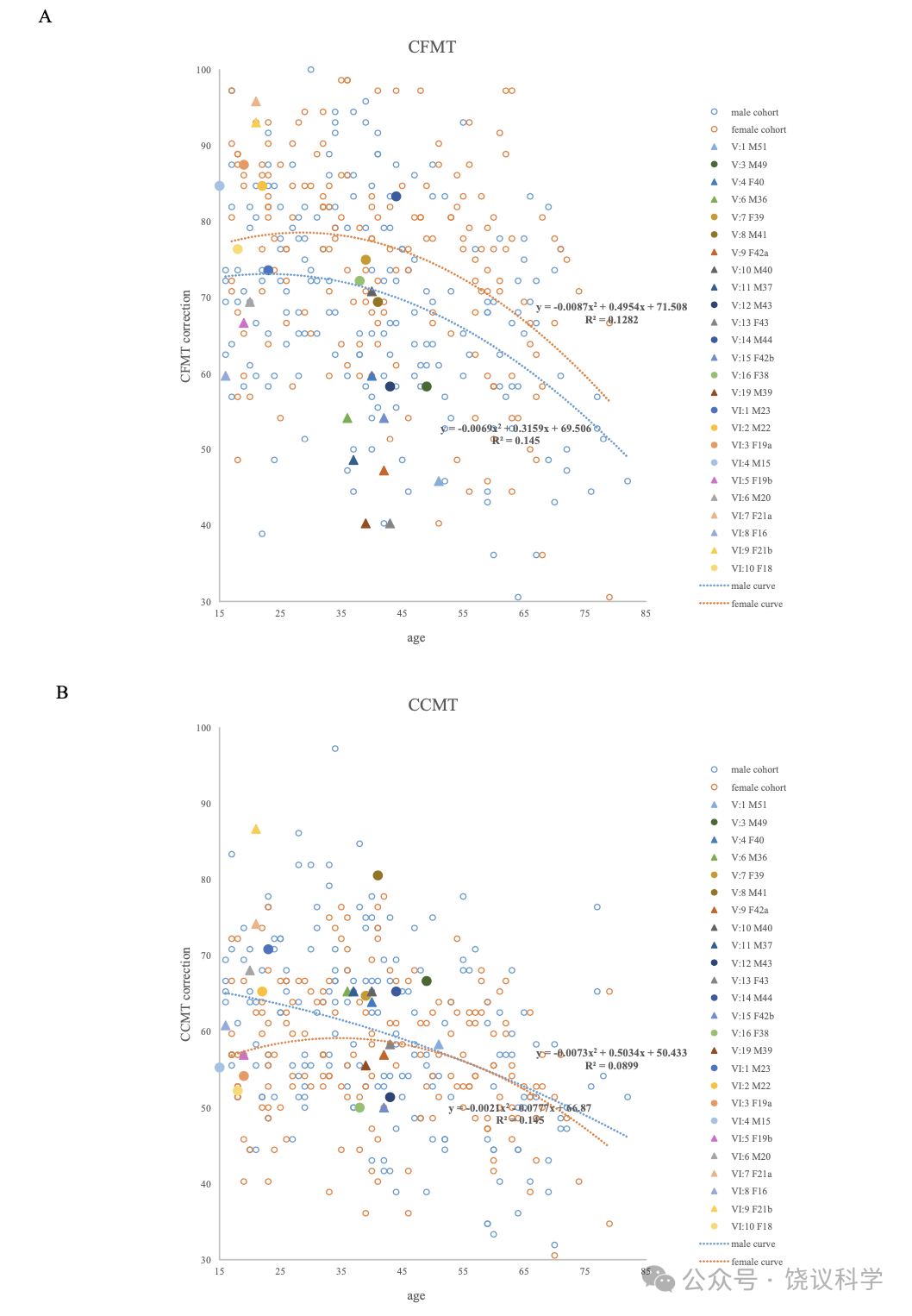
��ͼS1��Scatterplots of Age against Total Correct for��a��the CFMT-C and��b��the CCMT.
Male control individuals are shown as blue circles; female control individuals are shown as red circles; individuals with daily face recognition problems in Family A are shown as solid triangles; individuals with normal daily face recognition in Family A are shown as solid circles. The curves show the best fitting second-order polynomial relating age to the relevant performance measure with different genders, blue for male, red for female.
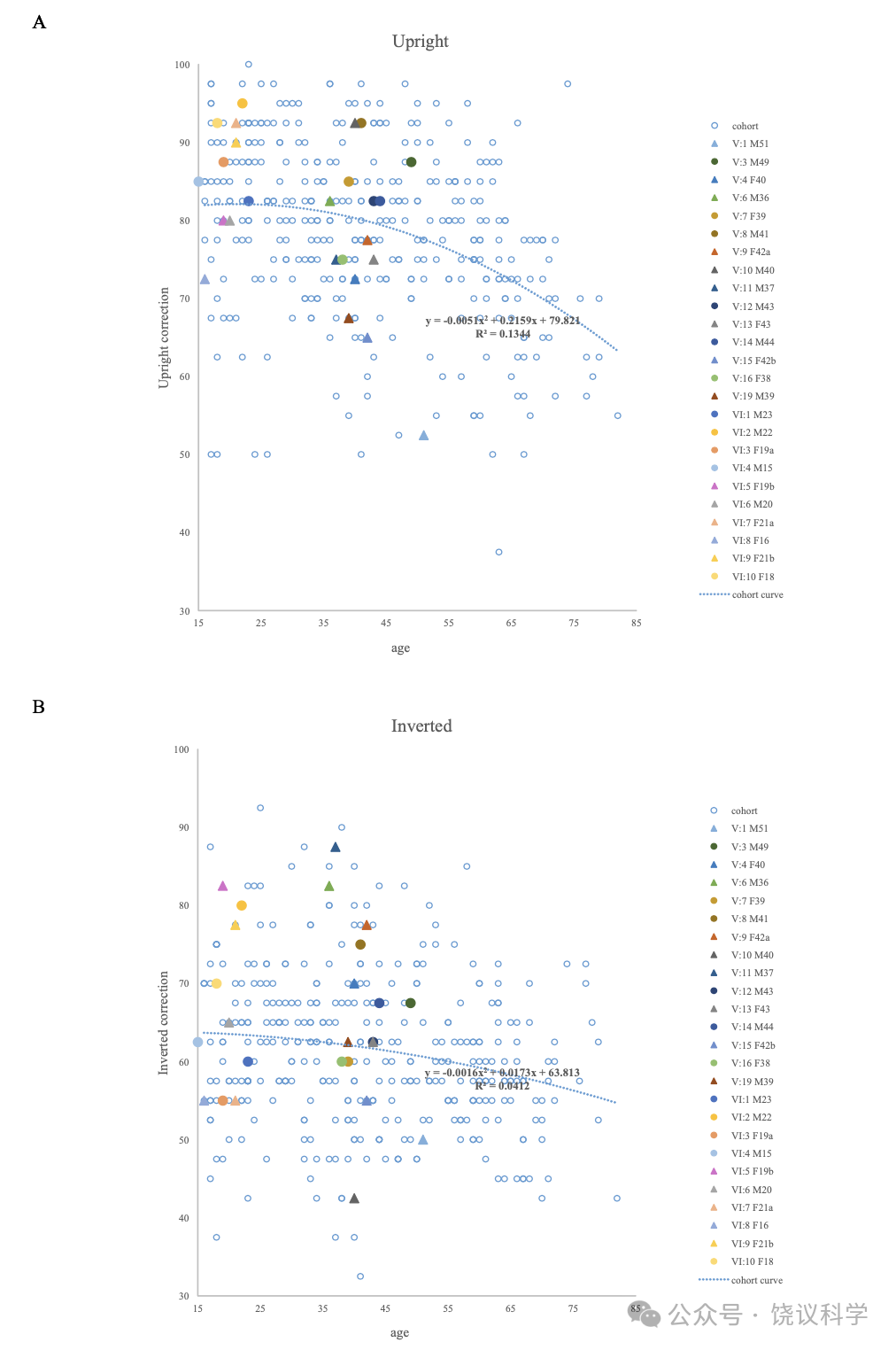
��ͼS2��Scatterplots of Age against Total Correct for��a��the Upright Face and��b��the Inverted Face.
Control individuals are shown as blue circles; individuals with daily face recognition problems in Family A are shown as solid triangles; individuals with normal daily face recognition in Family A are shown as solid circles. The curves show the best fitting second-order polynomial relating age to the relevant performance measure.
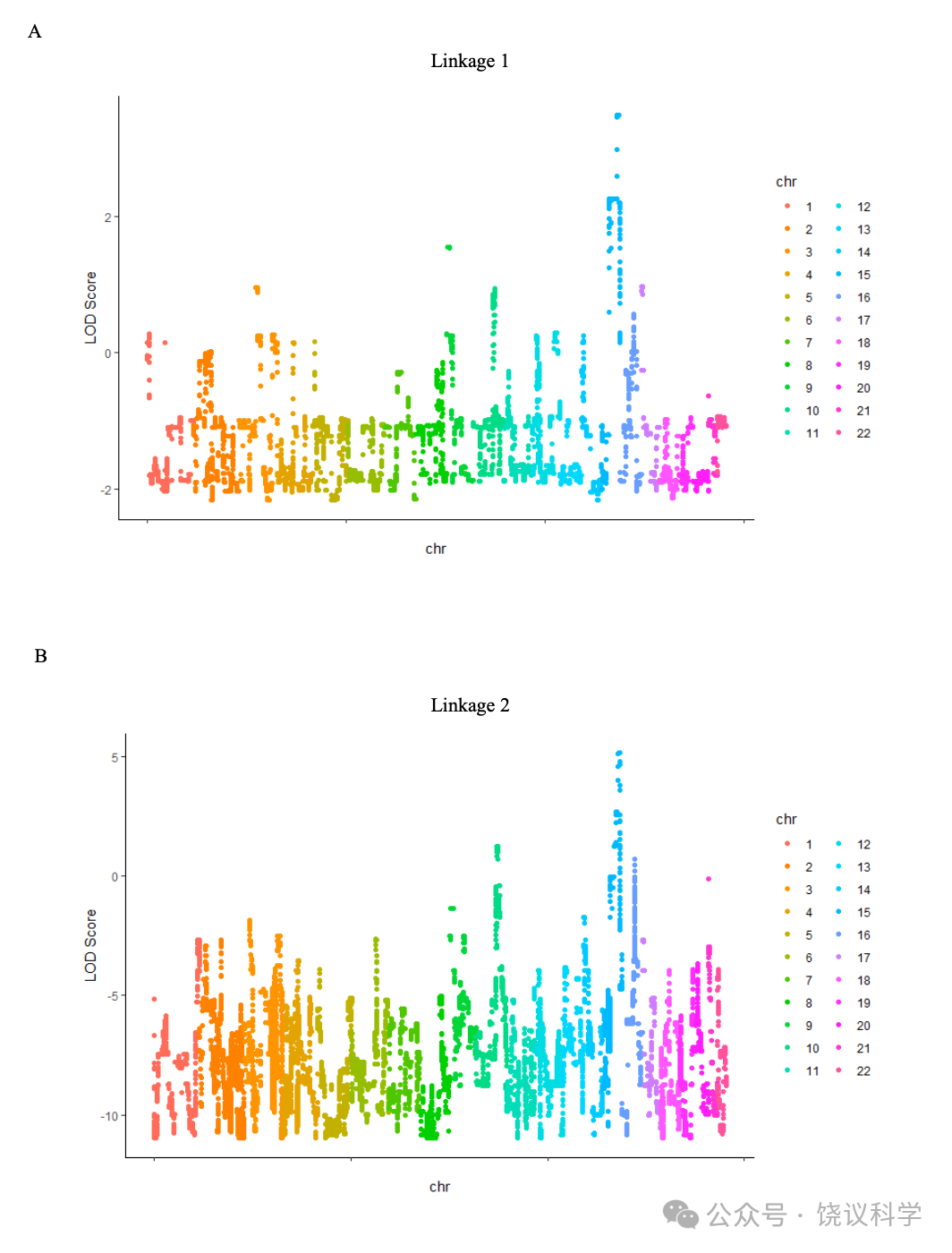
��ͼS3��Genome-wide Screen Results of CP-susceptibility Loci. Allele-sharing multipoint LOD scores are shown����a��for linkage 1, and��b��for linkage 2.
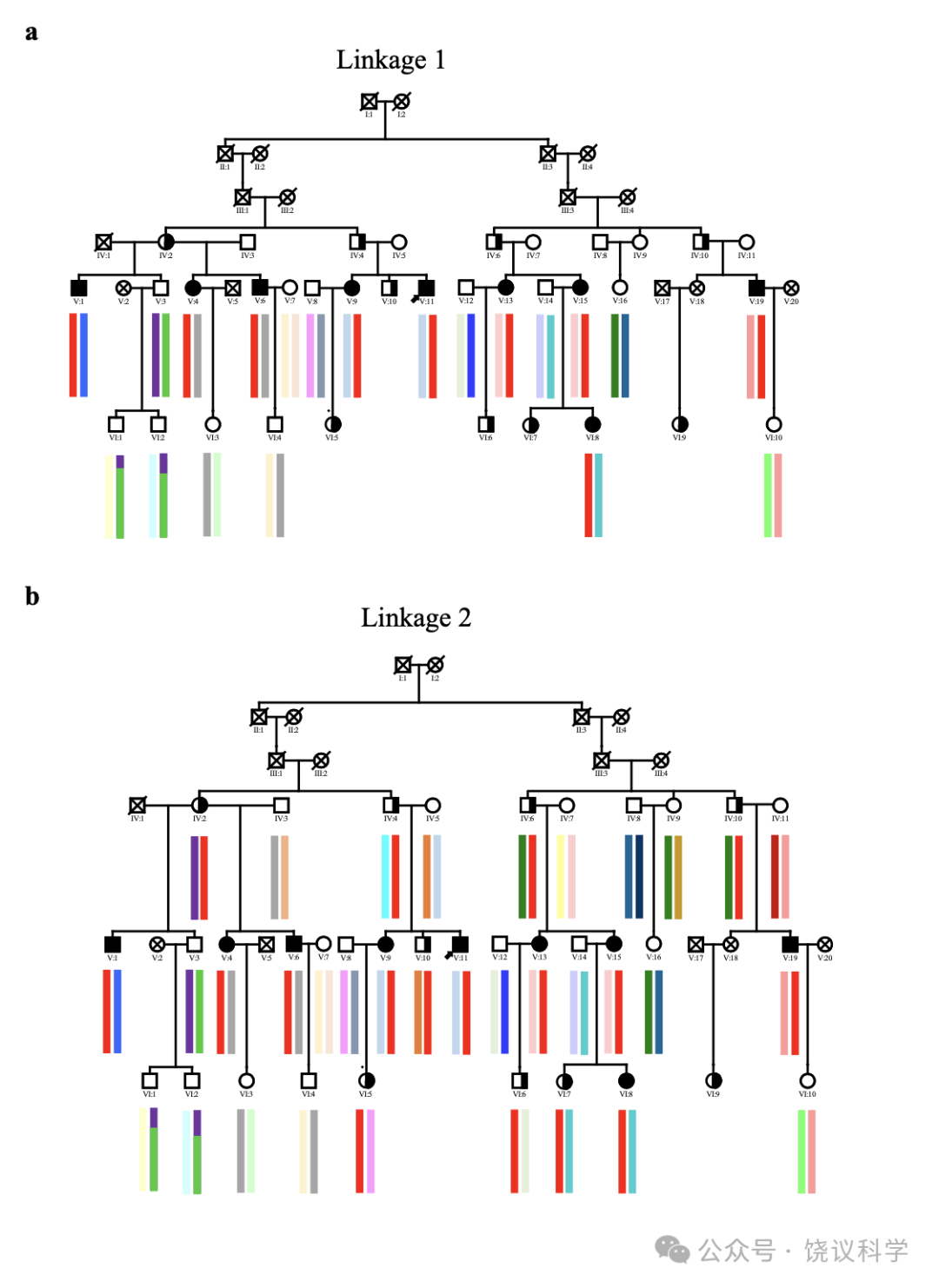
��ͼS4. Haplotype Analysis for MCR on the Autosomal Dominant CP Locus to 15q26
��a��Genotyping was performed for 20 individuals in linkage 1. Haplotypes for the 3.9 Mb MCR are color-coded, with red indicating co-segregating with CP.
��b��Genotyping was performed for 34 individuals in linkage 2. Haplotypes for the 2.64 Mb MCR are color-coded, with red indicating co-segregating with CP.
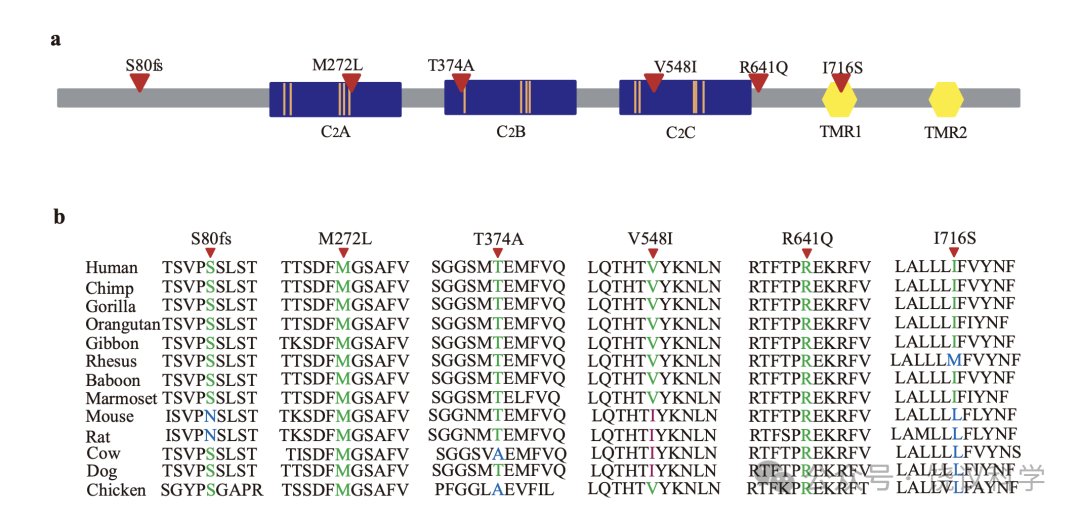
��ͼS5��The Positions in the MCTP2 Protein Corresponding to MCTP2 Mutations in CPs
��a��The domain plot of the MCTP2 protein and locations of alterations found in CPs. The blue rectangles represent C2 domains, the yellow hexagons represent transmembrane segments; the orange lines indicate putative Ca2+ binding sites; the red triangles indicate amino acid changes.
��b��Evolutionary conservation of amino acid residues in the MCTP2 protein in different species. Amino acids corresponding to mutations found in CP subjects are denoted in different colors. Green��identical to those in the human MCTP2, blue��non-identical to those in human MCTP2, fuchsia��non-identical amino acids but the same as the mutated in human MCTP2. Mutated amino acid residues are indicated by triangles on the top.
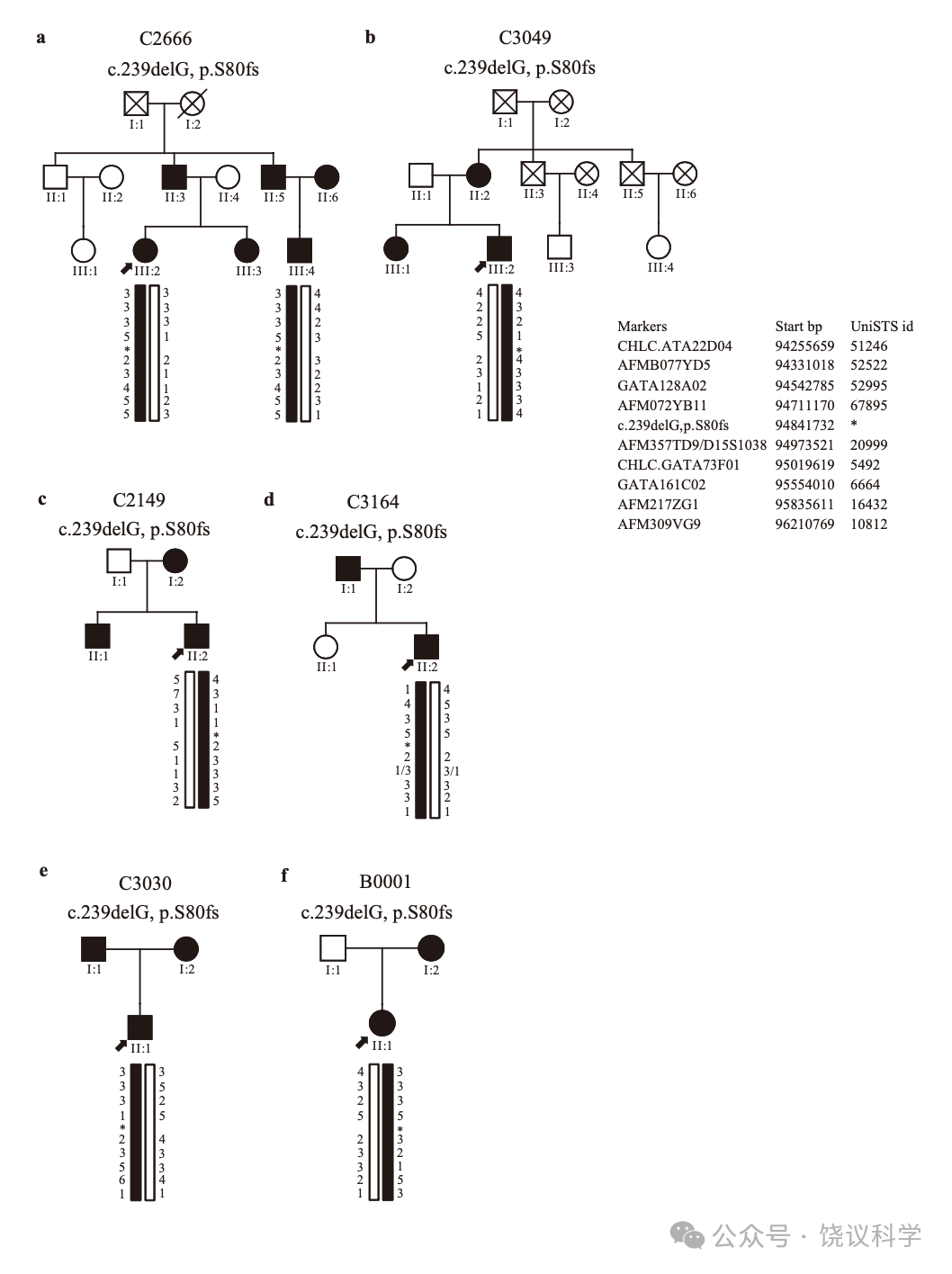
��ͼS6��Inferred Haplotypes around the c.239delG��p.S80fs��Mutation
These were estimated by comparison to microsatellites data collected on mother-father-offspring trios. Haplotypes coded in black contain the c.239delG��p.S80fs��mutation. The haplotype on the left is paternal and the haplotype on the right is maternal.
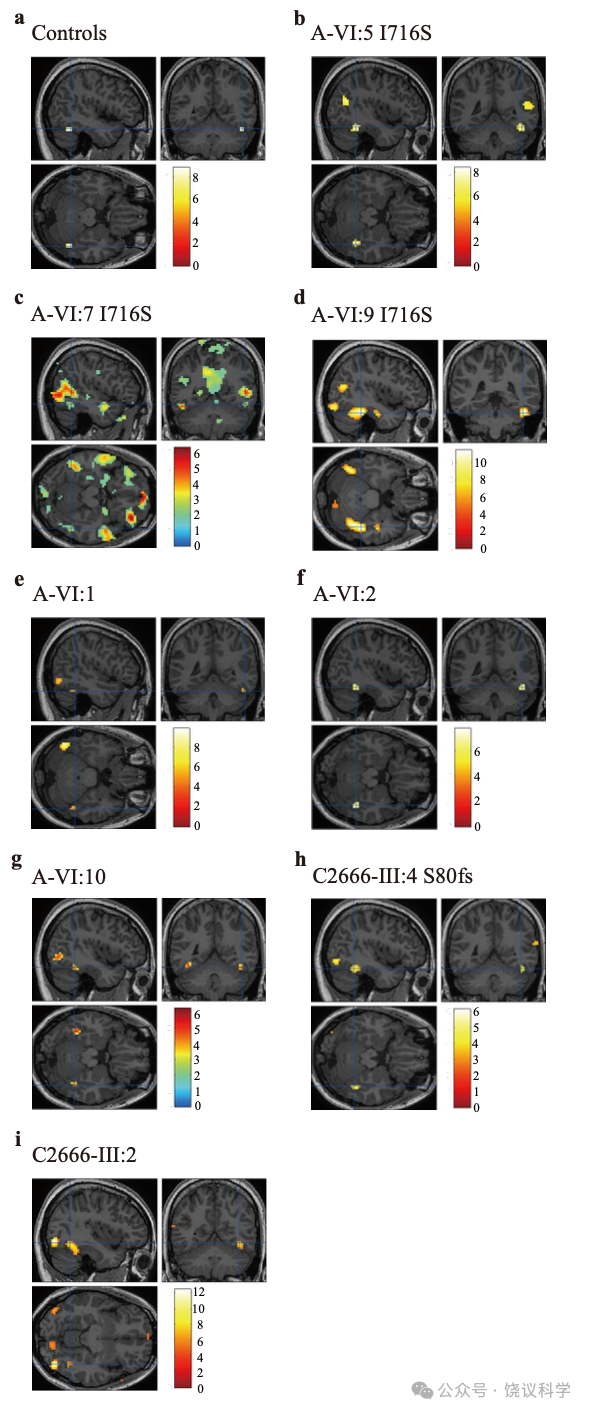
��ͼS7��Brain fMRI from the Localization Experiment��Identifying the FFA ROI.
Statistical parametric maps by contrasting the response to faces versus non-face stimuli, showing significant face-specific bilateral or unilateral activity in the FFA. Color scale represents statistical values of a one level t test comparing the fMRI signal while subjects viewed blocks of faces versus blocks of non-face stimuli.
��a��rFFA responses in normal controls, pFWE < 0.05.
��b��rFFA responses in A-VI:5 CP member with the I716S mutation, pFWE < 0.05.
��c��rFFA responses in A-VI:7 CP member with the I716S mutation, p < 0.05.
��d��rFFA responses in A-VI:9 CP member with the I716S mutation, pFWE < 0.05.
��e��rFFA responses in A-VI:1 non-CP member, pFWE < 0.05.
��f��rFFA responses in A-VI:2 non-CP member, pFWE < 0.05.
��g��rFFA responses in A-VI:10 non-CP member, p < 0.001.
��h��rFFA responses in C2666-III:4 S80fs CP member, p < 0.0001.
��i��rFFA responses in C2666-III:2 non-CP member, pFWE < 0.05.
��
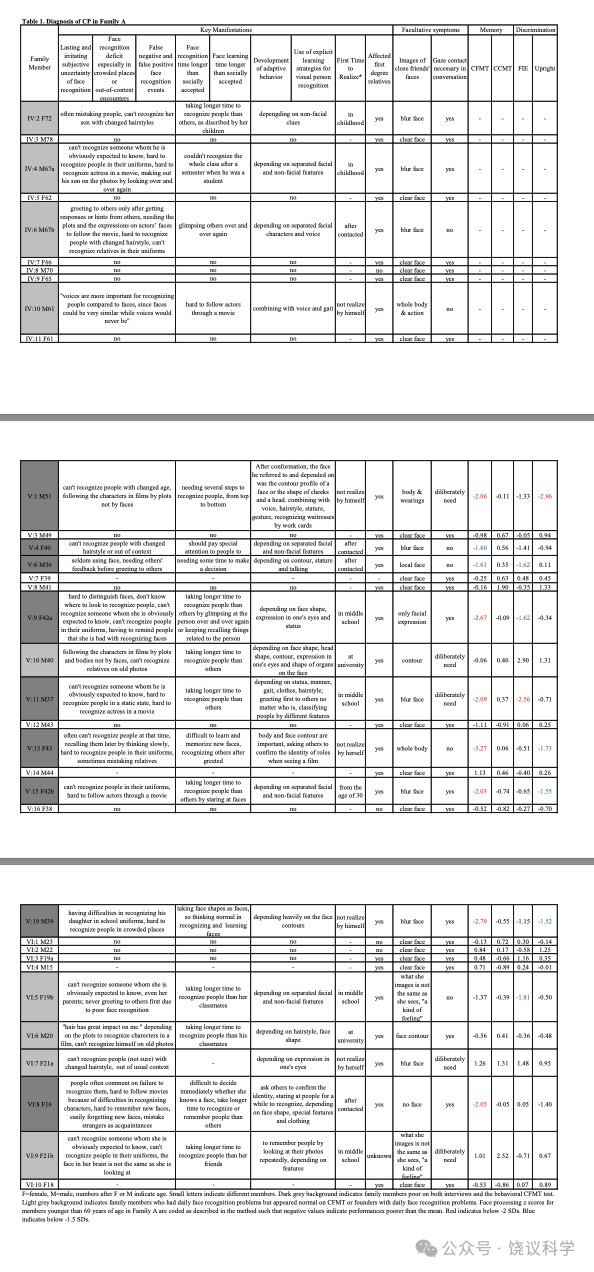
��1. Diagnosis of CP in Family A
F=female, M=male, numbers after F or M indicate age. Small letters indicate different members. Dark grey background indicates family members poor on both interviews and the behavioral CFMT test. Light grey background indicates family members who had daily face recognition problems but appeared normal on CFMT or founders with daily face recognition problems. Face processing Z scores for members younger than 60 years of age in Family A are coded as described in the method such that negative values indicate performances poorer than the mean. Red indicates below -2 SDs. Blue indicates below -1.5 SDs.
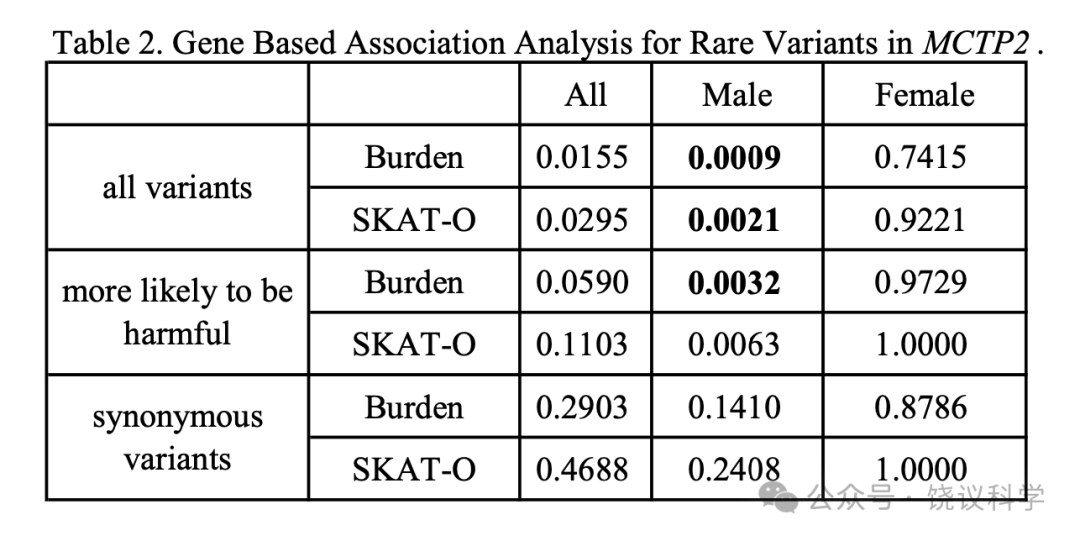
��2. Gene Based Association Analysis for Rare Variants in MCTP2
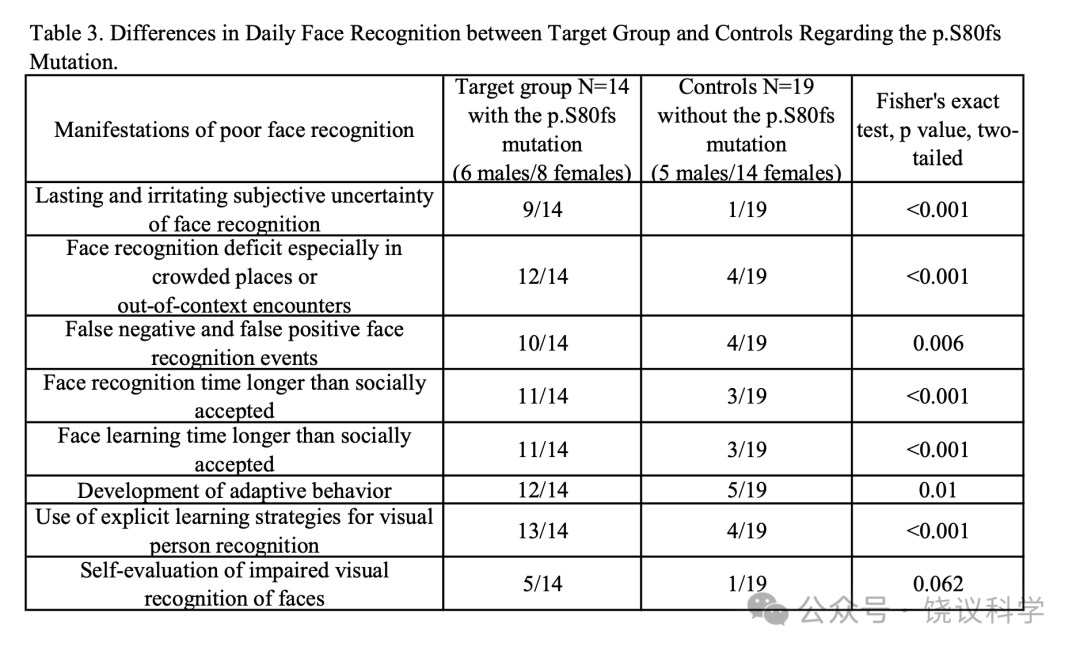
��3. Differences in Daily Face Recognition between Target Group and Controls Regarding the p.S80fs Mutation.
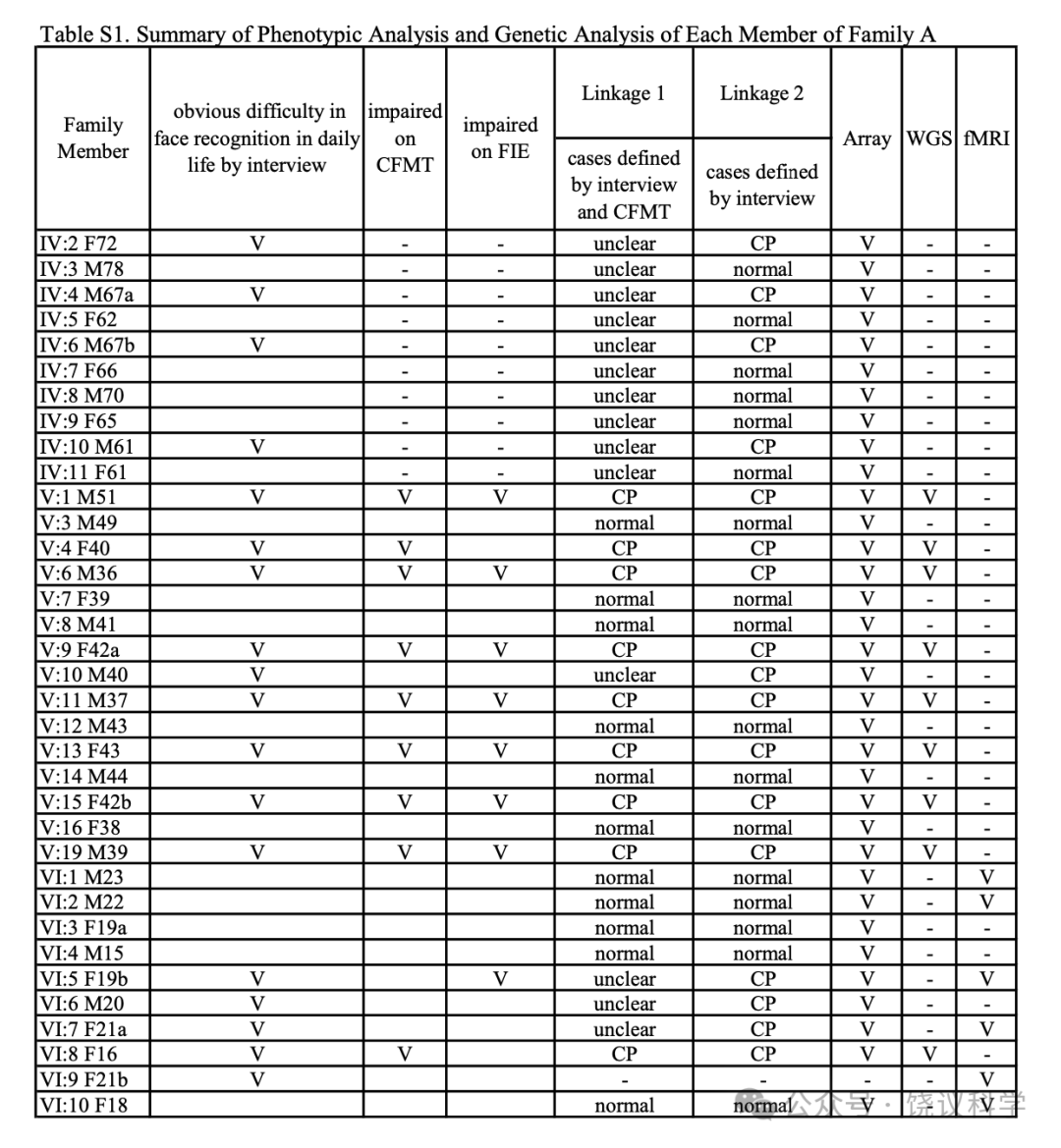
����S1. Summary of Phenotypic Analysis and Genetic Analysis of Each Member of Family A
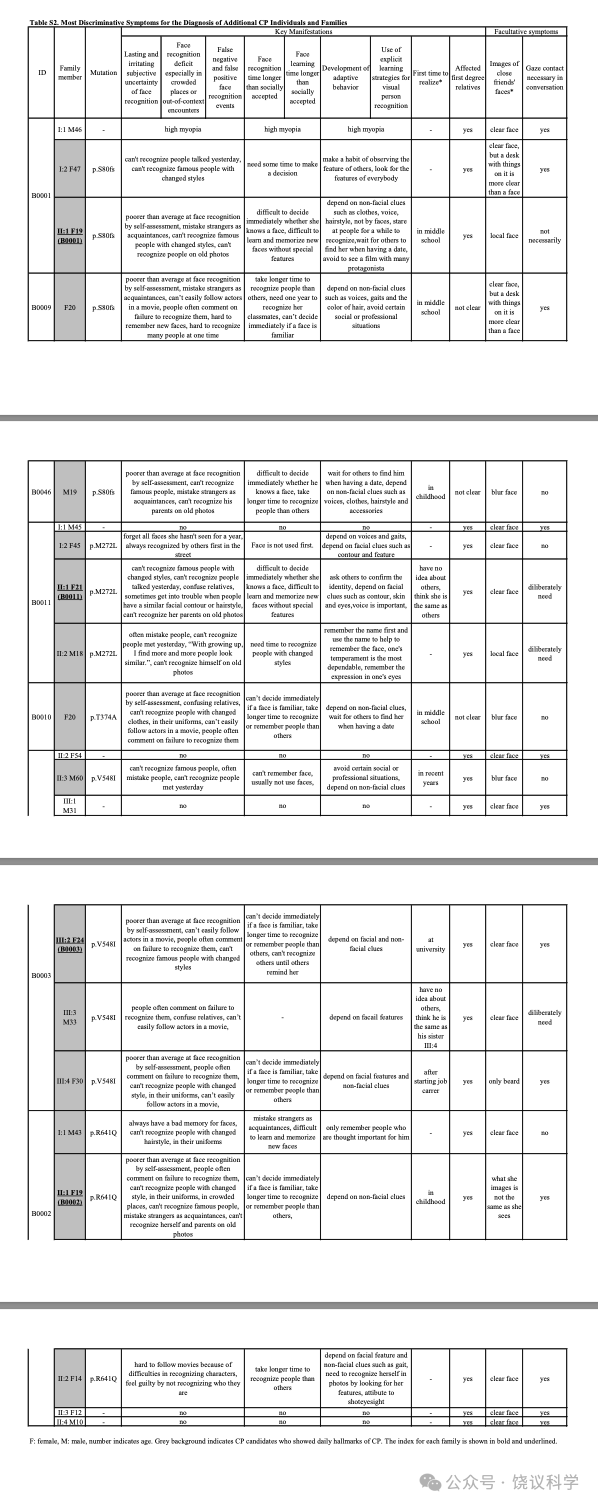
����S2. Most Discriminative Symptoms for the Diagnosis of Additional CP Individuals and Families
F��female, M��male, number indicates age. Grey background indicates CP candidates who showed daily hallmarks of CP. The index for each family is shown in bold and underlined.
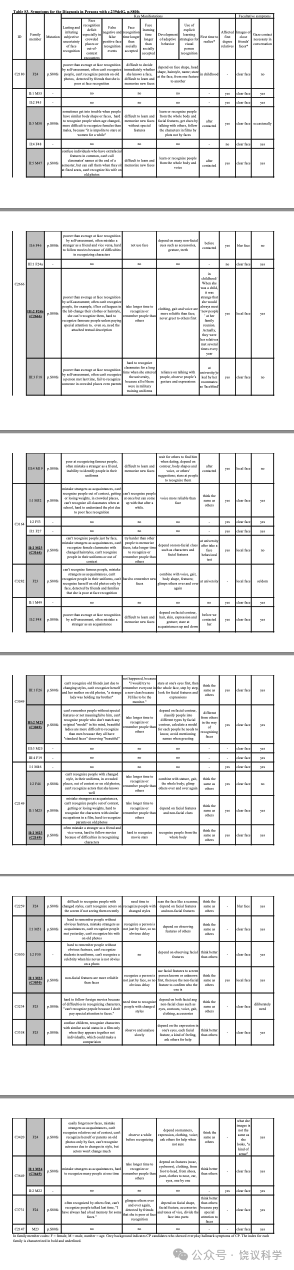
����S3. Symptoms for the Diagnosis in Persons with c.239delG��p.S80fs��
F��female, M��male, number indicates age. Grey background indicates CP candidates who showed everyday hallmark symptoms of CP. The index for each family is characterized in bold and underlined.
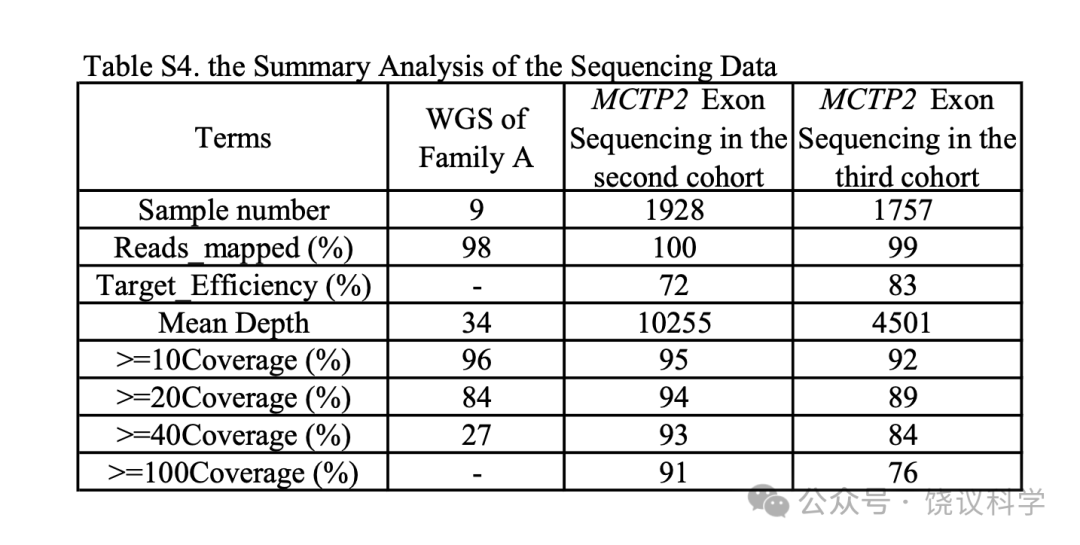
����S4. the Summary Analysis of the Sequencing Data
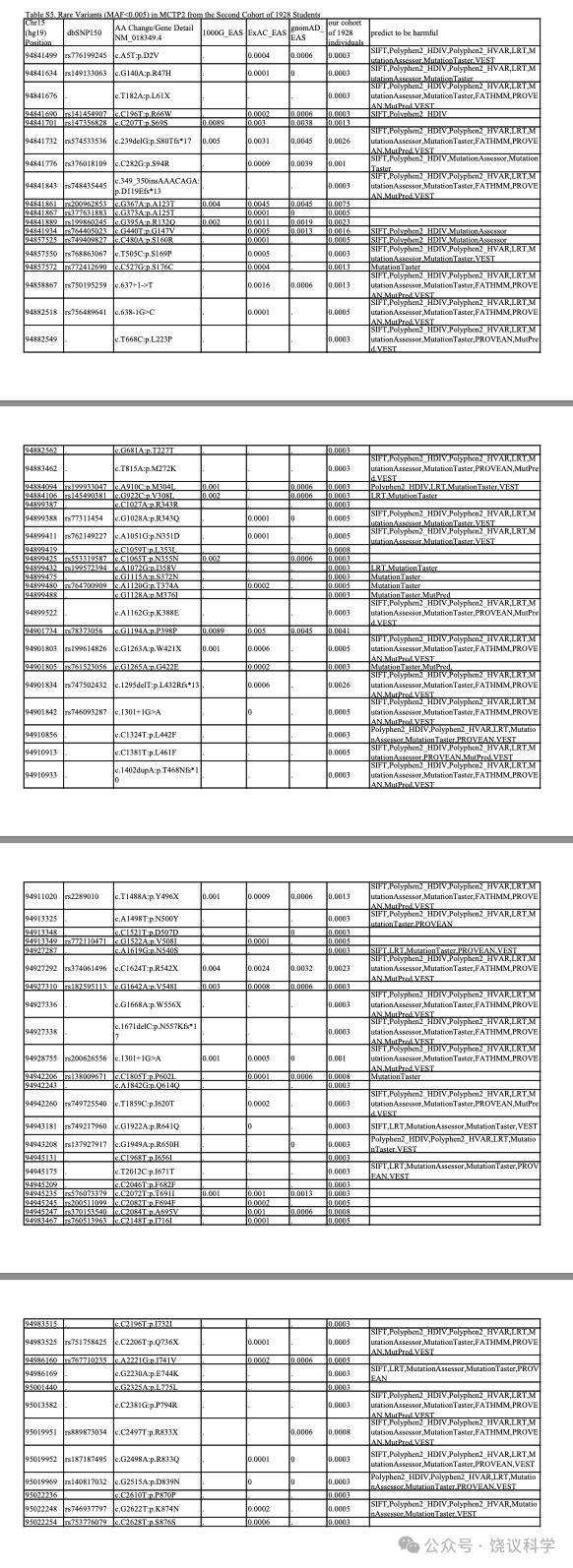
����S5. Rare Variants��MAF < 0.005��in MCTP2 from a Second Cohort of 1928 Students
* Start Position Based on the MCTP2 RefSeq��hg19, GenBank��NM_018349��; # dbSNP build 150

����S6. PCR Primer Sequences for Each Exon and Mutation Site of MCTP2
L, the left primer; R, the right primer; seq, the primer for sequencing
����S7. Candidate Genes Annotated by the NCBI Database��NC_000015.9��in MCR
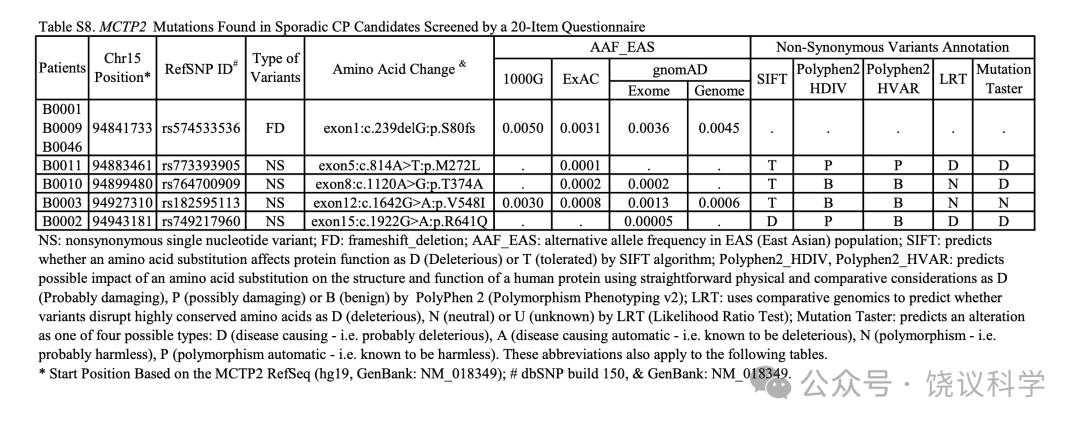
����S8. MCTP2 Mutations Found in Sporadic CP Candidates Screened by a 20-Item Questionnaire
NS��nonsynonymous single nucleotide variant; FD��frameshift_deletion; AAF_EAS��alternative allele frequency in EAS��East Asian��population; SIFT��predicts whether an amino acid substitution affects protein function as D��Deleterious��or T��tolerated��by SIFT algorithm; Polyphen2_HDIV, Polyphen2_HVAR��predicts possible impact of an amino acid substitution on the structure and function of a human protein using straightforward physical and comparative considerations as D��Probably damaging��, P��possibly damaging��or B��benign��byPolyPhen 2��Polymorphism Phenotyping v2��; LRT��uses comparative genomics to predict whether variants disrupt highly conserved amino acids as D��deleterious��, N��neutral��or U��unknown��by LRT��Likelihood Ratio Test��; Mutation Taster��predicts an alteration as one of four possible types��D��disease causing - i.e. probably deleterious��, A��disease causing automatic - i.e. known to be deleterious��, N��polymorphism - i.e. probably harmless��, P��polymorphism automatic - i.e. known to be harmless��. These abbreviations also apply to the following tables. * Start Position Based on the MCTP2 RefSeq��hg19, GenBank��NM_018349��; # dbSNP build 150, & GenBank��NM_018349.