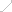������ǿ����һ�ָ��ϵ���Ԫ�������Կ��ƹؼ�ϸ�����ݻ���ı��������ļ��ˮƽ����֯������ת¼���Ӻ���ӣ����н��帴������Ը�Ƶ����Ŀ����������ӷ�����ϵ�������������ǿ�Ӱ��������ɵ���Ԫ������Ŀǰ�в������ЩԪ���ڼ���Ŀ�����������Ƿ���в�ͬ�����á���ǰ����Cell�Ϸ�����һƪ��Ϊ"Super-enhancers include classical enhancers and facilitators to fully activate gene expression"�������У�����ͨ���ؽ���Դ�ԵĶಿ�֦�-�鵰�׳�����ǿ�ӣ�չʾ������������������Ϣѧ�ϵ�Ч���ڹ����ϲ�ͬ��Ԫ�����ͣ�������ǿ�Ӻʹٽ���Ԫ����
�ٽ��ӱ���û����ǿ�ӻ��ԣ�Ȼ��������ȱʧ������£�������ǿ������ȫ�ϵ���л���ı��û�дٽ��ӣ�������ǿ�ӵ��н�����ļ����ǿ��RNAת¼����ǿ������������þ����١��ٽ��ӿ��Ի���ʹ�ã������������ڶಿ����ǿ���е�λ�ã����ڹ��ܲ㼶����ˣ��ٽ�������ǿ������ǿ�ӵĻ��Ժ�ȷ���л����ǿ��������Ҫ���á�

ͼƬժҪ
����
������ǿ�ӣ�super-enhancer ,SE����һ�ָ��ϵ���Ԫ�������Կ��ƹؼ�ϸ�����ݻ���ı�������ɾ�����ǿ�Ӻʹٽ�����ɣ���ͬ�������á��о����֣��ٽ��Ӷ��ھ�����ǿ�ӵĻ���������Ҫ����ǿ���ã�ȷ����Ŀ��������Ч���
���ܶԳ�����ǿ�ӽ����˹㷺�ķ����������ǵ�����������Ȼ�������Ŀǰ���о��������飬��ȷ��������ǿ���Ƕ����ľ�����ǿ��Ⱥ�廹�ǰ�����ͬ����Ԫ�����͵ĺ����塣�����Ŵ�����SEs�������ѣ����ѽ�SE���صĻ������仯��ϸ����ϵ�ͷֻ�����ر仯���ֿ�������ˣ���Ҫ���ϸ�ķ�����ȷ��������ǿ�ӵ���ʵ���ʡ�
С���-�鵰�׳�����ǿ�ӣ���-globin SE����-SE���������Ԫ�����ɵģ��������ظ��Ħ�-�鵰����λ��һ���ض��������ں�ϸ���ֻ��������ϵ�������֮ǰ���о��Ѿ��Ԧ�-SE�����˽��������������������ܹ������ϵ��������ľ�����ǿ�ӣ��Լ�����û����ǿ�ӹ��ܵ�Ԫ����Ȼ������Щ�о���û��̽��ÿ��Ԫ���ij�����Լ����Ԫ��֮��Ĺ��ܹ�ϵ��
����ͨ�����̸���С���-�鵰�׳�����ǿ�ӣ��ؽ�����ṹ����ʾ�����еĸ��ӹ�ϵ���о����֣�������ǿ���ɾ�����ǿ�Ӻʹٽ�����ɣ��ٽ��Ӷ�����ǿ������ǿ�ӵĻ�������Ҫ���á�
ʵ����
��С���-�鵰�׳�����ǿ�ӽ��й��̸��죬��Ϊ������ǿ��Ԫ���IJ���ƽ̨
��ͳ�ı༭�����ڵ�����λ�����н��ж������ͻ��Ĺ��̸����ʱ�Ҹ��ӣ���Ҫ���������ȷ����ȷ�ԣ������⡰��Ŀ��ЧӦ��������ܻ�Ӱ��ϸ���������Ѻͷֻ����Լ����ɺ�����С��ģ�͵�������
Ϊ�˽����Щ���⣬���߲��������¿����Ĵ���DNAƬ�ε�ȫ����װ��������Ʋ��ϳ��˰���Ұ���ͦ�-�鵰����-TAD�������Ʊ����86 kb��λ����ͨ������ø�鵼�Ļ����������������Щ��λ�������ϵ��Ѿ�ɾ����������-�鵰������λ��mESCs�У�ͼ1A����ʹ�����߿�����ÿ���ºϳɵĻ�����λ�Ͻ��л����������ͨ������ֻ��ͺ�ϸ������ϵͳ�����߶���Щ��ͬ�ĺ�ϸ�������˷�����

ͼ1 ��������ǿ�ӣ�����-SE��������С��ģ�ͺͽ���R2������С��ģ�ͣ��Բ���R2��ǿ��Ԫ���ij����
��û��������ǿ�ӵ�����£����������������
��ɾ��������ǿ�ӵ�����£�����-SE������Դ��EB�ĺ�ϸ��������ȫʧȥ�˦�-�鵰�ı��ɥʧ�ʳ���99.9%��������SEԪ����ص�����Ⱦɫ�ʱ�Ƕ���ʧ��ͼ1B��1C����û����ǿ��ʱ������������λ�ϵ�H3K27Ac������ȫ��ʧ��ֻ������̥��-�鵰������ص�һ���dz�С�ķ���Ȼ���ڣ�ͼ1B��������-SEģ��Ϊ�о�SEԪ���ں�ϸ�����ɹ����е������ṩ��һ����ȷ�Ļ���
����һ����ǿ�������Ħ�-�鵰��������R2-only�����-�鵰�ױ���������µ�����̥�������
Ϊ��ȷ��R2��ǿ�ӶԦ�-�鵰�ױ���ĸ��幱�ף����߽�����ǿ�ӻ�����������-SE�������R2�Ļ����������˱Ƚϣ�ͼ1A��������R2�ĺ�ϸ������Ħ�-�鵰��ΪԤ�ڵ�10%����Ԥ��ֵ����5����ͼ1C����
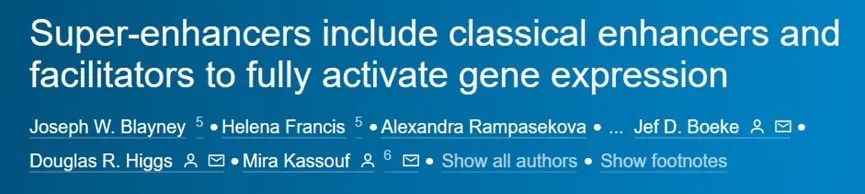
Ϊ�˽�һ���о�����R2�ı��ͣ�����������һ������R2��С��ģ�ͣ��������Դ�Ԧ�-SE������ǰ�Ħ�R2С����ȣ�����R2С��Ĵ���ʽϵͣ�ͼ1D��������R2С����������ͽ�С�����Լ�ƶѪ������Ƣ���״������ͼ2A����ֻ��һ��������С�������7��ʱ������������Щ�������R2��ǿ����С��������ѪҺѧ������������Ҫ���á�
ͼ2 ����R2��С����ֳ����ؼ����Ħ�-�鵰�ױ�����������
���߷���R2-only������С�����������ƶѪ��֢״���������ȥ��R1��R3��Rm��R4���ڦ�-�鵰�ı�������˱�Ԥ�ڸ����Ӱ�졣Ϊ���ų��������صĸ��ţ����߽�����һϵ�е�ʵ��ͷ��������ȣ�����ͨ��ӫ�⼤��ϸ����ѡ�����Ժ�ϸ��Ⱥ������˷����������ʾR2-only��ϸ��Ⱥ��ķֻ�״̬��WTͬ���dz����ơ��������ϸ�����ɺ�ϸ��Ⱥ����첻�ǵ���ƶѪ����Ҫԭ��Ϊ�˽�һ��ȷ�Ϧ�-�鵰�ı�����������߽�����RT-qPCR��RNA-seqʵ�顣
���˾��ȵ��ǣ���Ԥ�ڵ�50%��ȣ�R2-only��ϸ����������15%�Ħ�-�鵰�ף�ͼ2B���������ȥ����R1��R3��Rm��R4���ڦ�-�鵰�ı�����������ĸ���Ӱ�졣���۲쵽��-�鵰���������ε���������ı����µ���ͼ2C��������SEԪ��ȱʧ������£�R2Ԫ������������ǿ�����ԣ���δ����ļ��ˮƽ�Ĺ�����Ԫ��
������SEԪ��ȱʧ������£�R2Ԫ������������ǿ�����ԣ���δ����ļ��ˮƽ�Ĺ�������
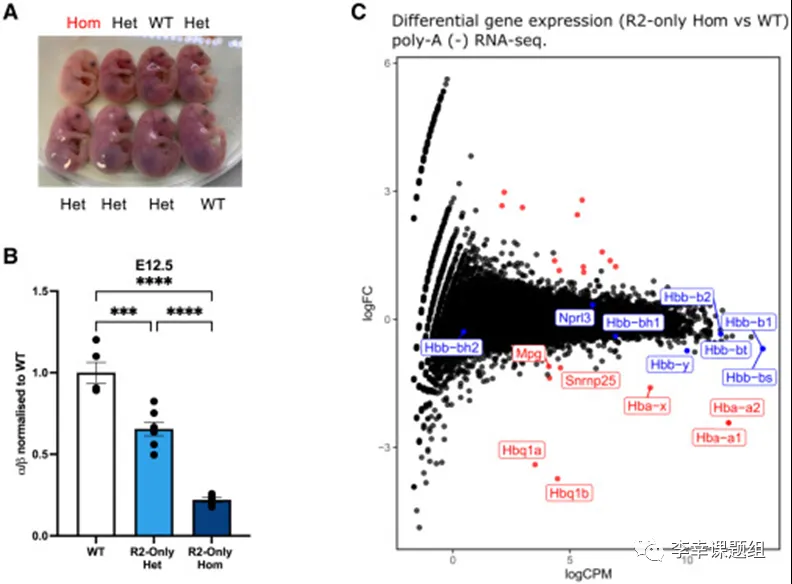
����ȥ��R1��R3��Rm��R4�����߶�R2-only ��-�鵰�����������������о������߷��֣���R2-only FL��ϸ���У�R2�ͦ�-�鵰����������Ȼ���ֿɼ��ԣ�����R1��R3��Rm��R4�ǻ����鷶Χ�ڱ仯��������ͼ3A����ͨ��ChIPmentationʵ�飬���߹۲쵽R2�ͦ�-�鵰���������ϴ��ڻ�����ǿ�Ӻ���������ص��鵰�����α�ǣ�������R2-onlyģ������Щ��ǵ�ˮƽ�������𡣴��⣬���߷���R2�ܹ���ļ��ϸ��������ת¼���ӣ��γɿ��ŵ�Ⱦɫ������ͼ3B��������ȱ�٦�-SE�е�����Ԫ����R2��Ȼ����������Ϊ��ǿ�ӵ����ԣ���ļת¼���Ӳ��������ŵ�Ⱦɫ�ʻ�����
Ϊ���о�R2��ȱ��������-SE�ɷֵ��������ļ�������ӵ����������߽�����ChIPmentationʵ�顣�����ʾ����R2-only FL��ϸ���У���ļ�н��帴���Mediator Complex����ԱMed1�ͺ�����ṹ��bromodomain���ĵ����� Brd4��ˮƽ���Խ��ͣ�ͼ3B�������⣬�����۲쵽R2-only��ϸ���У���-�鵰����������Pol II��ռλҲ�������٣�ͼ3B������û��������-SE�ɷֵ�����£�R2�Ĺ���������ļ�����ܵ�������Ӱ�졣
ͼ3 ��R2-only��ϸ���У�R2�����˻�����ǿ�ӵ����������������ӵ���ļ��������
��û��R1��R3��Rm��R4������£�R2��eRNAת¼��������
��ǿ��ͨ������л���ת¼��������ͬ���ȵ�˫��ת¼�����о����֣�R2-onlyϸ����ֻ��R2��ǿ����ʾ��ת¼����������-SE�ɷ���û�С������R2��ת¼�����ϱ�������ǿ�ӵ��������������ɷֵ�ȱʧ������ת¼�ļ��١�����ת¼����ǿ��ǿ��֮��Ĺ�ϵ�Լ�eRNA�Ƿ���й��ܵ�������Ȼ�������
Ϊ�˶����Ƚ�WT��R2-onlyϸ���е�R2 eRNAת¼���������ͨ�����С�����qPCR��ʵ�飬���߷�����R2-onlyϸ���У���WT��ȣ�R2 eRNA��ת¼��������Լ3����ͼ4A����
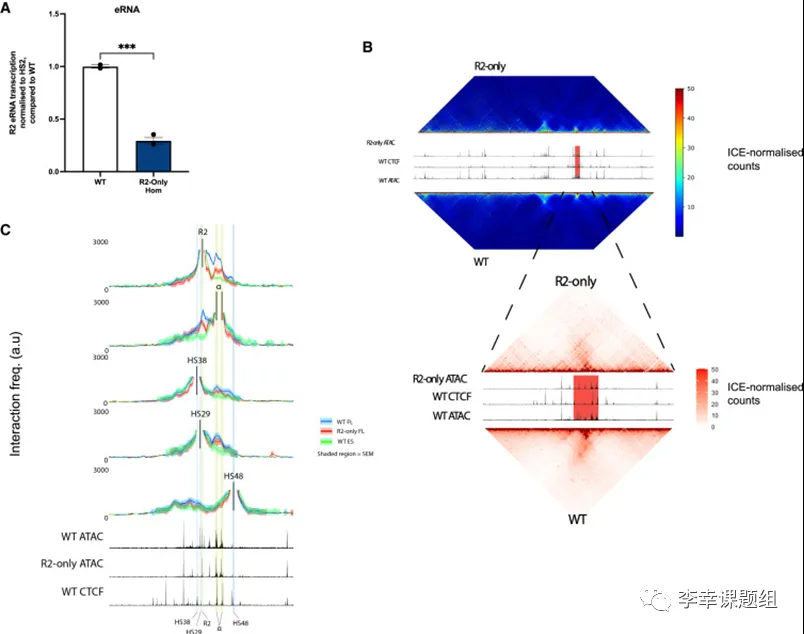
ͼ4 ��R2-only��ϸ���У�R2 eRNA��ת¼��R2-�������ӵ������Ƶ�ʶ�������
��ȱ��������-SE�ɷֵ�����£�R2-only����λ���е���ǿ��-������������ܵ���Ӱ��
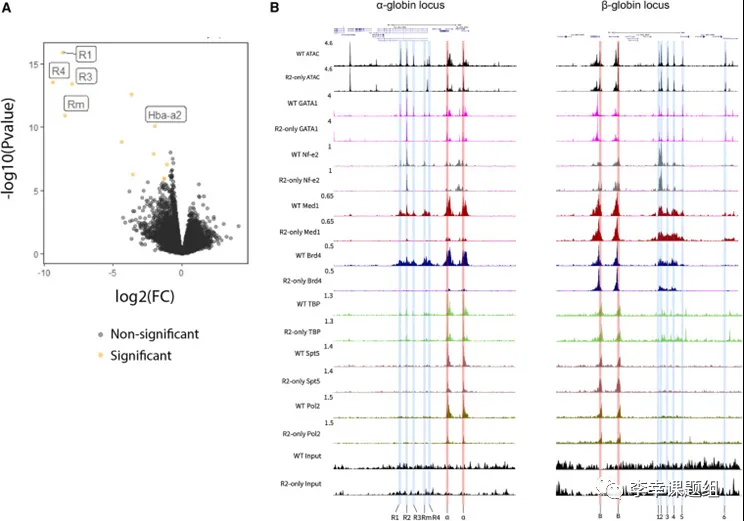
�����о���������ǿ����Ŀ�����֮����ڸ�Ƶ�ʵ�����á�Ȼ������Щ����õĻ��ƺ������Ĺ�ϵ�Բ��������ǰ���о����֣��ں�ϸ���У��ر���R1��R2�ɷ֣���-��ǿ�Ӿ������-�鵰�������ӷ�������á�Ϊ�˽�һ���о���R2-only����λ����R2���-�鵰�������ӵ��������������߲�����һ�ָ߷ֱ��ʵ�Ⱦɫ�ʹ���chromatin conformation capture ,3C������������Χ�Ʀ�-�鵰��3.3 Mb������λ������˱Ƚϡ�
Ⱦɫ���������ͼ��ʾ����R2-only FL��ϸ���У��о����֦�-�鵰����-TAD��Ⱦɫ�������Ƶ���������ͣ�ͼ4B��������CTCFλ������ΧDNA������ñ��ֲ��䣬��R2���-�鵰��������֮��������Ƶ���������ͣ�ͼ4C������������WT mESC������ȣ�R2-only FL��ϸ���е������Ƶ����Ȼ�����ϸߡ��������R2Ԫ���ڵ��ڦ�-�鵰�������������Ҫ���ã���������Ԫ����ȱʧ��δ��ȫ��������õķ�����
��-SE����ɲ��ֲ�����ȣ����������ֲ�ͬ�Ĺ���
��R2-onlyС��ģ���У�R2��ǿ����Ȼ�����˻�Ծ��ǿ�ӵ�������������ļ�������ӡ���Ŀ���������������á�����˫��eRNAת¼�����Լ��ϵ���-�鵰�ױ�������������������Ϊ��ȷ������SEԪ�������ã�����ʹ���������mESC��ϸ���ֻ�ϵͳ�����˲�ͬ���õĦ�-SE�ؽ���
���ߵij��������Ǧ�-SE����ɾ��е���ЧӦ������ͨ����������SE�����ɾ��Ԫ����ʵ��ó��ġ�Ϊ��������֤��Щ���֣������ڰ����mESCs���ؽ�����ͬ��ɾ���������ʾ��ɾ��R1��R2���������˦�-�鵰�ı����ɾ��R3��Rm��R4��Ӱ�첻���ԣ�ͼ5A��5C������Щ�����һ��֧��������֮ǰ�ķ��֡�
���߽�һ���о��˽���-SE�ĵڶ�����Ҫ������R1���²��뵽����ǿ�ӵĦ���-SEλ���е�Ч����Ȼ������ʹͬʱ����������Ҫ�����ӵ�ģ�ͣ�R1R2-only����Ҳ���ָ���ˮƽ�Ħ�-�鵰��ת¼��ͼ5A����
���߷��֣�����R3��Rm��R4Ԫ������û�д�ͳ��ǿ�ӵĻ��ԣ��������ƺ���Ȼ�������Ħ�-SE����������Ҫ�����߽����Ϊ���ٽ��ӡ���Ϊ���о�R3��Rm��R4�����R1��R2����ã�����������һ������ǿ�ӵζ�ϵ�С������ؽ���ԭʼ�Ħ�-SE��������������R3/Rm/R4��������ϣ�ͼ5B����
����Ϊÿ��ģ�������˶����¡����ͨ��PCR��Sanger������֤�����ǵ������ԡ�ʹ��JASPAR��Sasquatch�ȼ��㹤�߽�����Ԥ�⣬ȷ������Ƶ�ģ��û������ش�������DZ�ڹ��ܵ����С�ͨ��ATAC-seqʵ��֤������ÿ��ģ���У�����ӦԪ����ص�Ⱦɫ���ں�ϸ���б�ÿɼ��ԣ���������λ��Ŀɼ���û������ĸı䣨ͼ5C����
�������²����˴ٽ��ӣ�R3��Rm��R4����������R1��R2�Ļ����У�������������ǿ������ǿ�ӵ������������ʾ����R3Ԫ�����²��뵽R1R2-only�����н���ʹ�����������Լ10%����ͳ��ѧ�����ԣ��������²���Rm��R4���и�������Ч�����ֱ�ʹR1R2-only�ı���������50%��80%�����⣬���²���R4Ԫ��������H3K27Ac��R1��R2Ԫ���ϵ��������ӣ�����R4����Ҫ�����Ǵٽ�����������ǿ�ӵ���ȫ���ԣ�ͼ5D����
��R3Ԫ�����²���ʱ�����۴���Rm��R4�����ߣ�����ʹ��-�鵰��ת¼�ϵ���������ͬ�ij̶ȡ�Ȼ�����ڰ���R4�Ļ���������²���Rm��Ч����С����֮��Ȼ����ˣ�Rm��R4�ڴٽ�������ǿ��R1��R2���ܷ����ƺ��в����ص�������R4��Ч����Rm��ǿ��

ͼ5 Ϊ�˷�����ȫ��DZ����R1��R2��ǿ��������R3��Rm��R4Ԫ��
R4�ľ�ԮDZ��ȡ������λ�õ��ض���
Ϊ��̽��R4���и�ǿ��ԮDZ����ԭ���������·�����DNase-seq���ݼ������������-SEԪ�������˻�������������ʾ��R1��R2��������ת¼���ӻ�����ӵ�DNase�㼣�źš�Ȼ������Rm��R4��ȣ�R3���и���ĺ�ϸ��ת¼���ӻ�����Ҳ�õ���DNase�㼣�źŵ�֧�֣�ͼ6A������Ȼ���ܴ�������δ֪�ӵIJ��룬�����ߵ����ݱ�����R3��Rm��R4�ľ�Ԯ������������ȡ����������ļת¼���ӵ�������
R3��Rm��R4�ľ�Ԯ�������������-�鵰��������֮��ľ���ʸ���ء�ͨ�����²���R4����ͬλ�õ�ģ���У����߷���R4�ľ�Ԯ����������ȡ���������У����������������ӵľ���������أ�ͼ6B����
Ϊ�˲���Ԫ��λ�õ���Ҫ�ԣ�������R1R2-onlyģ���в�����R3������ʾ���ٲ���λ��Ŀɼ�Ⱦɫ�ʡ������ʾ����R3�ƶ���������-�鵰�������ӵ�λ�öԻ���������������Ӱ�죬�����R1R2-onlyģ����R3����ԭʼλ���ϵľ�ԮЧ��ԼΪ12%����R1R2R3[R4]ģ���л�����������˳���85%��ͼ6C������ǿ�ұ���R4��λ�ã������������У������ھ�Ԯ������﷽���Ч�����ž��������á�
R2-only FL��ϸ���У�R2���-�鵰��������֮��������Ƶ�ʽ��͡����߲²�R4�����Ӧ�-SE��������֮��������Ƶ�ʷ�����ܷ������á�Ϊ�˲����Ƿ�ͨ������R2��������֮������Ծ�����Ծ�ԮR2-only��ת¼ȱ�ݣ����߽�R2���뵽R4��λ�ã������˾��ȵ��ǣ���R2���������Ӳ�û�жԻ�������������Ӱ�죨ͼ6D���������������ͨ�������ϵ����Խӽ�����ȫ�ָ�R2�Ļ��ԡ�

ͼ6 R3��Rm��R4����Ի�����Ҫ�����ǵ�λ��������
�������ಿ����ǿ����ʶ��ٽ���
����̽���������ಿ����ǿ�Ӵ��Ƿ����붨��Ĵٽ������Ƶ�Ԫ����ͨ���Ԧ�-�鵰��LCR�е�HS1Ԫ�������о������־�����ȱ�����ڵ���ǿ�ӻ��ԣ����ڦ�-�鵰�������ϱ��ֳ��˴ٽ��ӵ�������ʵ������ʾ����HS1�����ڦ�-�鵰�״ٽ��ӵ�λ�ã����-�鵰�״ٽ���һ��������ʾ����R1R2��λ�������������DZ�����۲쵽��-�鵰�ױ����36%���ӣ�ͼ6E�������ܱ�ԭ���ٽ��ӵľ���Ч���ϵͣ�RmԼ50%��R4 >80%���������о�Ϊ��һ���˽�ٽ��ӵ������ṩ����Ҫ������
����
�Դ�1981���״�������ǿ��Ԫ������������ǿ�ӵ��о��Ѿ��dz��㷺��������ǿ�Ӵ��Ѿ����о�����ʮ���꣬��������Ȼ�����Ļ���ԭ���˽����ޡ��������͵���ǿ�Ӵر����������а���������ǿ�ӵȡ�����ѡ���о�������ǿ������Ϊ���Ǿ�����ȷ����㷺Ӧ�õ��ص㡣������ˣ�������Ȼ��ʶ����ͬ����ǿ�Ӵؿ����в�ͬ�����û��ơ�
������ǿ�ӣ�SE�����и�ˮƽ��H3K27Ac���Ρ�ת¼���Ӻ͵����ӽ�ϣ��Լ��ɷ�֮�����Ļ�������롣�������������о�֤��SE�ܹ������ˮƽ�Ļ�������Ŀǰ����SE����ͨ��ǿ�Ӵ�֮��Ĺ��������Բ�������о����֣�SE�����Բ�ͬ�ķ�ʽ��ϣ��е��Ǽӷ�ʽ�ģ��е��ǷǼӷ�ʽ�ġ�Ȼ���������о��е����ƣ���SE�Ľ����о��Բ���ȫ�����⣬SE��Ŀ������Ӱ������ڲ�ͬ������֮���������졣
�������о��У����߶Ԧ�-�鵰�׳�����ǿ�ӽ�����ȫ��Ľ��ʣ����о�������ɷ�֮��ĺ�����ʽ���о����֣����е�һ���ɷ�R2�����ܵ���ʵ����ǿ�ӵ�����DZ��������Ҫ�����ĸ��ɷֵ���ϡ�����R2�����˻�����ǿ�ӵ����������䵥�����ò����ܴﵽ��ˮƽ�Ħ�-�鵰�ױ��
���¹�����-�鵰�׳�����ǿ�ӽ�ʾ������ɷ�֮��ĸ��Ӻ�����ϵ����ǰ�ĵ���ɾ��ģ������ȫ�������ֺ������о�������R1��R2����Эͬ���ã���R3��Rm��R4��ٽ���R1��R2�Ļ��
�о����֣��ڻ������п��ܴ������ƵĴٽ���Ԫ������MYC������ǿ�ӵ��о���ʾ�������ĸ��ɷ���HepG2ϸ����û�пɼ�����ǿ�ӻ��ԡ����Ƶأ������о�Ҳ���������������һЩԪ����Ȼ������ǿ�ӻ��Խ���������������ǿ�ӵĹ���ȴ������Ҫ����Щ���ֱ����ٽ��ӿ����dz�����ǿ�ӵ��ձ�������
Ŀǰ�в�����ٽ��������ǿ������ǿ�ӵĻ��ԡ��о��������ٽ��ӿ���ͨ���ṩת¼���ӻ�ͬ�ӵ���ļ���ȶ��Ե�Эͬ�������������á����⣬�ٽ��Ӻ���ǿ�ӵ��ʵ�λ�ÿ��ܻ����γ��ض�����ά�ṹ����Ȥ���ǣ��ٽ��ӵIJ�νṹ�����ȡ�������ǵ�λ�ö��������С�������о������֣��ڹ�Ӭ�д������Ƶ�Ԫ�������Dz�����ǿ�ӣ�����ͨ��������Ԫ����������ٽ�����֮�������á�
һЩ�о���������ǿ�Ӵأ�����������ǿ�ӣ�����ͨ��Эͬ�����γɺ��и�Ũ�ȵ���֯������ת¼���ӡ��������Ӻ�RNA�ۺ�øII�ľ۽�������Щ�Ǻ˽ṹ���γɻ���Ŀǰ�������飬��Һ-Һ������ת¼���Ӳ�������۱��������Щ���۶���Ҫ���ض�����ά�ռ�����ļ�㹻������ת¼���ӡ��о����֣�ͨ������ת¼���ӽ��λ���ܶȣ�R4�ܹ��ָ���������R2����ʵ�֡�
�ܵ���˵�����ߵ��о����������������ǿ�ӿ����γɸ��ӵĵ���Ԫ�����磬չ�ֳ�Эͬ�������Эͬ��ǿ�Ĺ��ܡ����ߵ��о��ṩ��֤�ݣ�֧�ֳ�����ǿ���ɾ�����ǿ�Ӻʹٽ�������ö������������ԡ�