2024��ŵ��������ѧ��ҽѧ��������λ����microRNA�Ŀ�ѧ��Victor Ambros��Gary Ruvkun�����ŵ��ʹѧ��������ٴξ۽���microRNA��microRNA��Ϊת¼����ص���Ҫһ�����о�������ѧ���ܡ����ӻ��ơ������༲���о�����������Ҫ���塣
���ľ۽�����ϵͳ��microRNA�ķ�����ʷ�����½�չ��������̽�ָ�������δ���������ͷ�չ����

1
microRNA�ķ���
20����70�������ʡ������ѧ��Bob Horvitzʵ���Ҽ������˴����߳���Ŵ�ͻ�䡣���IJ�ʿ��Victor Ambros����߳�Ļ������˴����Ŵ�ѧ������������������Ҫ���ܵĻ���Lin-4��Lin-14��
1982�꣬��һλ��ʿ��Gary Ruvkun������Bob Horvitzʵ���ң�������Lin-4��Lin-14��̽����1993�꣬������Cell�Ϸ����˱����������£���ʾ��Lin-4������һ��СRNA������ת¼�����Lin-14����ĵ��ױ���ˮƽ[1-2]�����ǵ�һ�ν�ʾ���ַDZ���СRNA�鵼��ȫ�µĻ�����ط�ʽ��
���ǣ���һ������һֱ����������λ��ѧ�ң�Lin-4�鵼�ĵ����Ƿ����߳���еģ����Ǵ����������������������Ǹߵȶ������ڵ���Ҫ���ط�ʽ? 1993-2000��䣬Ambrosʵ�������˴�������ȥѰ��������Lin-4��СRNA������ͽ���������Լ�Ҳ��˶��뿪�˹����ѧ���������é˹ѧԺ������ʵ����Ҳ���ھ��Ѷ�ȱ����eBay�Ϲ�����������豸��ֱ��2000�꣬Ruvkunʵ���ҳɹ�����������һ��microRNA����ʷ�ϼ�Ϊ��Ҫ�ķ��ӣ�Let-7������һ��ʼLet-7Ҳ�����߳��з��ֵģ��������Ǻܿ췢����Let-7�����ϵı����ԣ����ڰ��������Ӭ���ڵ����������ж���������Let-7[3-4]��
�Ӵˣ�microRNA�ķ��ֺ��о���ʼ����ƽ�ʱ�ڣ��ɰ���ǧ��microRNA�ڲ�ͬ�����б����֡�Խ��Խ����о�������������ѧ�ϣ������Ǹߵȶ���ķ����ͷֻ��У������ŷdz���Ҫ�����á�
Ambros��2008��ع������ʷʱ���������ڵ�֪Let-7�ķ��ּ����ڽ����еı�����֮��ļ����������һ���ֵĻ�ʱ�����壺����2000��������Ҷ���Gary ���ǹ���Let-7������֮�������Ӵ���ʮ����ӣ����������Ҷ�����������֪�����ڣ���������������Lin-4��Let-7��һ�������й��ϵĵ��ط��Ӽ���ij�Ա����˶����п϶�����������������RNAؽ������[5]��
����After reading their paper in the autumn of 2000, I had to set aside 10 minutes to stare out the window and reorganize my view of the universe. Now we knew that lin-4 and let-7 RNAs were members of an evolutionarily ancient class of regulatory molecules, and so it was finally obvious that there must be other RNAs like them yet to be discovered in animals����
2
microRNA������ϵͳ
2.1 ���ֵ�ǰҹ
2000��Let-7�ķ��ֵ�ȼ�˿�ѧ�ҶԷ��ֺ��о�microRNA����Ȥ������microRNA�ڲ��鶯���з��Ӻε����ã��ڸ���ϵͳ����ε��ػ�������Ȼ��Ҫ�Ŵ�ѧ֤��[6]��
����ϵͳ�������Ǻ�������ϵͳ���Ǹߵȶ����������һ�����Ҿ�ϸ�ķ���ϵͳ��2003�����ң�����ѧ���ǿ�ʼע�microRNA������ϵͳ�п��ܾ߱���Ҫ�ĵ������ã�һЩʵ���ҿ�ʼ����Dicer �Լ�����microRNA�����ó�С�����ͬʱ���ڸ��������С����֯�ж�microRNA���м����ʾ������������ϸ���ͷ�����������[7]��һЩmicroRNA��������֯��Ҳ�и߱���[8-10]����Щ֤����ʾmicroRNA���ܷ�����Ҫ�ĵ������á�һЩʵ���Ҳ��ù����ع����߹��̻���ķ�ʽ����microRNA������С��������ϵͳ�й۲쵽���Ա���[11]�����ǣ���Щ����ؽ��ͨ����������microRNA�����ó�������С����֤ʵ��������ϵͳ�еĹ����Լ��о����Ƿ������õķ��ӻ��ơ�
2.2 �����о�
Dicer�Dz�������microRNA�Ĺؼ��ӹ�����[12-13]��Dicerȱʧ����ɾ������microRNA����������⣬�Ӷ�����ȱʧ�������о������������ó�Dicer�ᵼ����Ѫϵͳ�����Լ�B ��T�ܰ�ϸ�����ڷ�����������ȱ��[14-16]����ѧ�����Ʋⵥ��microRNA����Ҳ����������ϵͳ�з�����Ҫ���á�������һ�Ʋ⣬һЩʵ���ҿ�ʼ�����ض�microRNA�ó�С��ͬʱ����������и߱����microRNA����ת����С�����о���ЩmicroRNA�������Ƿ���дٰ����á��Ƿ����е�ʱ��������microRNA����ǿ�����أ�
Klaus Rajewsky��Tyler Jacks��Martin Turner��Carlo Croce��ʵ���Ҹ��ݽ����е����б����ԡ���֯�����ԡ��Լ������и߱������������Ŀ��Ͷ�����Ϊ������֪�ļ���������ѧ�Ͼ�����Ҫ���ܵ�microRNA������miR-17-92��miR-155�Լ�miR-150��
2.3 miR-17~92
miR-17~92������2004����Bϸ���ܰ���������Ʒ�м�������[17]����һ���ϴ��microRNA������壬����miR-17~92��miR-106a��363��miR-106b��25�����ڻ����鲻ͬλ�õ�ͬԴ����أ�����4��miRNA�Ǽ��壨miR-17��miR-18��miR-19��miR-92�Ǽ��壩������13��microRNA��ʵ��֤����Mycת����С����ͨ��������Ⱦ������miR-17~92���Լӿ��ܰ����γ�[18]��Rajewskyʵ���Ҳ�ʿ��Ф�������õ�ʱ�Ѿ�����cre-loxpϵͳ���������ܰ�ϸ������������miR-17~92ת����С��������Щת����С����Է�����Ƣ����ܰͽ��״��ܰ�ϸ�����ֳ������ϵͳ���ң��Ӷ�����ת����С����ǰ��������Щ�����ʾ��miR-17~92���ܰ�ϸ�����淢������Ҫ������[19]��
ͬһʱ�ڣ�Jacksʵ���Ҳ�ʿ��Andrea Ventura����������miR-17~92���������������ó�С����������ȫ���ó�������£�С����̥����������ȱ�ݣ��ó�С��Bϸ��������������ͬʱ�������о�����miR-17~92�����ó�С�������Ҫ������miR-17~92������ó���������������صĹ��ܽ���[20]��������ʵ����ͨ���о�miR-17~92�ó��������ܰ�ϸ������Bim��Pten��miR-17~92���ص���Ҫ�л�����miR-17~92�ó�ϸ����ɾ��Pten��Bim��һ���������Ժܴ�̶��ϻز�miR-17~92�ó����µı��͡�
2.4 miR-155
miR-155��һ�����ܰ������и߱����microRNA�������ض�����ϸ��Ⱥ�и߱���[9]��2007�꣬Rajewskyʵ���Һ�Bradleyʵ���ҹ���������miR-155�ó���ת����С����Science�ϱ�������������ƪ���½�ʾmiR -155������ϵͳ�з�����Ҫ���ã����������������ķ�Ӧ��������Ҫ����[21-22]���ó�miR-155������������Bϸ�����٣�������������������Bϸ�����ӡ��ó�miR-155Ҳ�����Tϸ���ֻ���ϸ�����ӷ���ȱ�ݡ�֮�ã�Turnerʵ���ҵ��о���һ����ʾ����Bϸ����miR-155��Ҫͨ������Pu.1���ƿ�������Ӧ�����[23]����Nussenzweigʵ��������ù��������ﺬ��miR-155���λ��ͻ���Aicdaת����С��֤����AicdaҲ��miR-155����Bϸ�����ܵ�һ����Ҫ�л���[24]��
2.5 miR-150
ͨ����С������ϵͳ�м��microRNA������[25]������miR-150��Ҫ�ڳ���ľ�Ϣ̬B��T�ܰ�ϸ���б�����Dz�����ǰ��ϸ���б�����ֽ������Եı���ģʽ������ζ��miR-150��B��T�ܰ�ϸ���з�������Ĺ��ܡ�ͨ������miR-150ȫ���ó�С���ת����С��Rajewskyʵ���Ҳ�ʿ��Ф��������miR-150��ȱʧ�ᵼ��B1ϸ�����࣬��������Ӧ����ǿ����miR-150ת����С����ᵼ���ܰ�ϸ����������[26]�����ӻ����о�����miR-150��Ҫͨ������ת¼����c-Myb�ĵ��ױ�����ʵ���书�ܵģ��������Ŵ�ѧˮƽ��֤��c-Myb�Ӻ�С����c-Myb����ˮƽ�����Ϳ�����ɸ�miR-150ת����С�����Ƶı��͡�
��������microRNA���ں������о��У�����ѧ��ͨ�������ó�С���о�ȷ����miR-142��miR-146a����Ӧ�����ߵ�������Ҫ����[27-28]��ͨ��������microRNA�ó�����ת����С����о�����Щʵ����ȷ����microRNA�鵼��ת¼������ڸߵȶ��������ѧ��������������ϵͳ�з�������Ҫ�����á�
ͬʱ��ͨ����microRNA��ϰл����Ԥ������Ӻϻ��ߴ��ϰл����ó�С��ΪmicroRNA��ε��ذл����Լ���ε�������ѧ�����ṩ�˹ؼ����Ŵ�ѧ֤�ݣ�������microRNA�ڲ�ͬϸ����������ξ����ذл������ķ��ӻ��ƾ����ش����塣����ѧ�һ�ͨ������microRNA��mRNA�������Լ�CHIP-seq��̽����microRNA���������ص��Լ�����ܵ����ء�2010���һƪ�о������ܰ�ϸ�������microRNA����ת¼�Լ�ת¼��ˮƽ���ܵ����ܵĵ��أ�Ҳ����˵��microRNA���������ڻ��������ص���Ҫһ��[29]��
���ǣ������ڵ�ʱ�ļ����ֶΣ�microRNA��ϰл���ȱ�������ͷ���ˮƽ�ϵ�ʵ����֤������microRNA�Դ���DZ�ڰл�����mRNA�͵���ˮƽ�ϵĵ���Ҳȱ������������ҲӦ��ע����������о��У�һЩ�������졢�߷�ȡ���Ԥ��Ϊ�й��ܵ�microRNA�ں�����С���Ŵ�ѧ�о��б�����û�����Ե�����ѧ���ܣ����߹����������Ա��ͣ������ó�û��������Թ����쳣������miR-181[30]��miR-210[31]�Լ�miR-148[32-33]��Ф����ʵ���ҹ�����miR-148a�Լ�ͬԴmicroRNA��miR-148b��miR-152���ó���ת����С����ЩС����з����������ܹ������ƻ���Bϸ����������[33]���ó�miR-148���microRNA������ϵͳ��û�з������Ա��ͣ�ʵ������δ����������ˣ�С���Ŵ�ѧ֤����Ȼ��microRNA�����о��Ľ������microRNA�о�����Ҫ���塣
3
microRNA���ػ���֮��
����Խ��Խ�����ҪmicroRNA�ķ��ֺ��о�������microRNA���ذл������ķ��ӻ��Ƴ�ΪmicroRNA�о�����Ľ�����ȵ㡣
2009�꣬David Bartel�������������ܽ���microRNA ʶ����л���ķ��ӻ��ƣ�������microRNA seed region��ƥ�䣬UTR���������������Ӱ�죬microRNA�Լ������mRNA��ϸ����Ũ�ȵ����ز���������Щ�������Ӱ��microRNA�л�������Ƴ̶�[34]��������ΪmicroRNA����ͨ��ͬʱ�Ժܶ�mRNA�ı�����������Ӷ�ʵ�ֶ�����ѧ���̵ĵ��ء�Ҳ����˵�����ٸ��л������ˮƽ��С�仯����ʵ��microRNA��ǿ���ܡ�
ͬ�꣬Ф������RajewskyҲ����һƪ����microRNA������ϵͳ�е��ػ��Ƶ�����[35]������microRNA���ͨ��������л�������������ѧ��������ˡ��ؼ��л�����һ���ۡ�����С���Ŵ�ѧ֤�ݣ���������ˣ�һ��microRNA�л���ĵ�����ϸ�����������ԣ��ض�ϸ���ڵ�������ػ�Ӱ��microRNA�л���mRNA��ʶ��͵��ء�������ͬ�л�����в�ͬ�ļ��������ԡ�microRNA��ͨ����Ϊ������Ĺؼ�������б�������ĵ�����ʵ��������ѧ���ܡ�����һЩ�о���������������ѧ����������Ago2 CLIP�������죬�Լ�Խ��Խ����Ŵ�ѧ֤�ݱ�����microRNA������ѧ������ͨ���Թؼ��л���ĵ�����ʵ�ֵġ���ѧ���Ƿ��֣��ڲ�ͬ����ϸ�������У�microRNA��Ҫͨ����Ϊ������ļ����л�����б���ˮƽ�ϵĵ�����ʵ���书�ܵġ�����miR-155����Bϸ������Ҫ��ͨ������Pu.1, Aicda�Լ�Smad5�����ӹ��ܣ���Tϸ��������Ҫͨ������Socs-1, Sp1r1�Լ�c-maf�����ӹ���[36]��2018�꣬Rudenskyʵ����������ѧ�����о���miR-155�ڲ�ͬ����ϸ���н�ϵİл���mRNA�Լ��л���mRNA����ˮƽ�ĵ��أ��Ӷ��ڷ���ˮƽ��֤��ϸ�����������Զ�microRNA��ϰл���mRNA�͵������DZ����Ӱ��[37]��
��һ����Ҫ��microRNA��miR-17~92��Rajewsky��Ф����ʵ���Ҷ����ϵͳ���о�Ҳ�ṩ�˺ܶ���Ҫ�ļ��⡣�ڷ��ӻ����ϣ����ܺܶ�microRNA��Ҫ���ڰл���mRNA�Ľ��⣬miR-17~92����Ҫ���ذл���ķ��롣���Զ�mRNA�͵��ױ���ˮƽ����ȷ����������microRNA���ػ����dz���Ҫ��Ф����ʵ����������ϸ�����������ֻ����ó�������miR-17~92[19,38-42]������miR-17~92�ڲ�ͬ����ϸ���Լ����ǵIJ�ͬ�����ξ��в�ͬ�Ĺ��ܺؼ��л����ң�miR-17~92��Bϸ���е��ó����������ʵ�ֶԲ�ͬ�л���ĵ��ء��ڹ���ˮƽ�ϣ���Bϸ�����ó�miR-17~92�ᵼ�¿���Ӧ��ȱ�ݣ������������γ��ܰ��� [43]��2019�꣬Rajewskyʵ���ұ�����miR-17��92�ؼ��л���Bim����ҪmiR-17��92���λ��ͻ��С��[44]�����о��Ӱл���Ƕ�̽����microRNA���ص�ϸ�����������Ժؼ��л����microRNA���ܵĹ��ס�Bim������miR-17��92���λ��ͻ��С���ʾ��Bim�ı����������鵼miR-17��92���ܵ���Ҫ�ԣ����Խ��ʹ�miR-17��92�ó�С��ı��ͣ���Ҳ������Bϸ��������miR-17��92��Bim����ˮƽ�ĵ��ز������ԣ���Bϸ������ҲӰ�첻��ԭ��ܿ����������Bim����ˮƽ̫�ߣ�������miR-17��92���Ե��ڵ���ֵ[45]��
4
�ܽ�
microRNA��Ϊ���������������ص���Ҫһ������Ŀǰ���ֵ���Ҫ�DZ���RNA����Ԫ�����������������ķ�����RNA������Ϊ��Ҫ�Ŵ���Ϣ���ӵĹ������ŷdz���Ҫ�ļ�ֵ��
���Ŷ�microRNA����ѧ���ܺͷ��ӻ����о������룬��Խ��Խ��С��������Ŵ�ѧ֤�ݣ��о�������mRNA�����ϸ��Ӷ�����ĵ����Ѿ���Ϊ����������ѧ����һ�ؼ��о������ڿ����¹������г�¶��ɵ�mRNA����Ҳչ����RNA�о���ת��Ӧ�÷�����DZ����mRNA�Ѿ���Ϊһ���ؼ��Ļ���������ƽ̨��mRNA�����˷dz����ӣ������������������Ρ��༭������ͽ��⣬��ŵ����������������ĸ����ԺͶ����ԡ�
��microRNAΪ������ͼչʾ��microRNA��ϵ��ذл���mRNA�������ܵ�����˳ʽ�ͷ�ʽ����Ԫ����Ӱ�죬����mRNA������5����3���Ƿ��������Լ�ϸ�����������һЩ����[46]��microRNA���ط�ʽ������Ҳ�dz����ӣ�����ͨ���ٽ��л���mRNA������������䷭����ʵ�ֵ��ء��������о���ΪmicroRNA���Դٽ�mRNA����ˮƽ�ϵ�[47]����ˣ�mRNA����ĵ��ػ��ƻ���Ҫ�������ϵͳ�Ե��о���
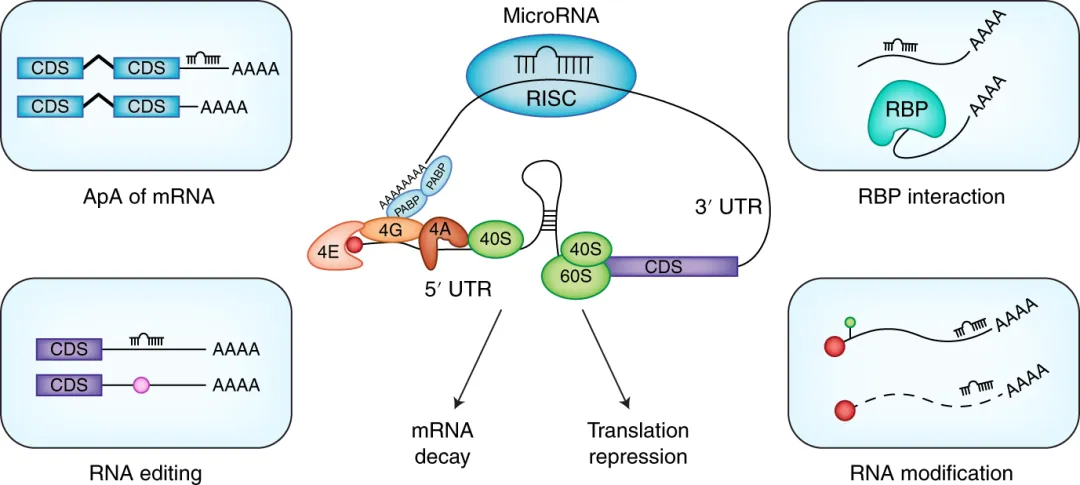
ͼƬ��Դ��Chen P, Liao K, Xiao C. Nat Immunol 19,
1040- 1042��2018��.[46]
��ϵ������ķ�չ�����������ϵͳ���о�microRNA���صķ�ʽ��
һ������RNA-seq��Ribo-Seq���Ͷ�������ȫ�����microRNA��mRNA����ˮƽ���������ʡ��͵���ˮƽ��Ӱ�졣
������Ұ���ͺ��ض�microRNA�ó�С���ԭ��ϸ��������Ago-CLIP����������microRNA��ϵİл���[48]��
��������CRISPR��ͨ��ɸѡ�Լ������İл�����й��ܷ������ҵ���microRNA�����й��İл���
�ģ�����������������ѧʵ�鷽������˳ʽ�ͷ�ʽ����ԭ������polyAβ����CLIP���λ�����й���λ�Һ�RISC�������л�����RNA��ϵ��ף� RNA�Ķ������������ṹ��������ϵͳ����ѧ˼ά����������microRNA���ذл����ģ�͡����⣬����microRNA���λ��ͻ��С����������Щ������ϵͳ�б�֤���к���Ҫ���ܵ�microRNA�ؼ��л���ͬʱ̽�������UTR��microRNA���λ���ڼ����������ǰ�֢���������߲��е�ͻ���Լ��²��Խϸߵĵ���������죨SNV��[49]���������С��ģ�͡���Щ�о����Ը��õش��Ŵ�������ѧ���о�microRNA�Ĺ��ܺ����û��ơ�
5
microRNA�о���չ��
5.1 �л���鵼��microRNA����
TDMD, Target-directed miRNA degradation
microRNA�����ǽ������·��ֵ�microRNA���ػ��ƣ���˵��ͨ������mRNA��trigger���յ�microRNA�γ�Ago-miRNA-trigger������Ӷ��Կ��Խ�ϵ�trigger mRNA����ӦmiRNA���λ���microRNA���н��⡣�û���������Zamore��Steitzʵ������2010�걨��[50-51]��2020��Bartel��Mendellʵ���ҽ�һ������������������ķ���øZSWIM8�ͷ��ػ�����;��[52-53]��Ŀǰ�û����Ѿ���֤���ڰ�����Ӭ��С��ȶ��ֶ������ڴ��ڣ���������ѧ��������û�����Ȼ�д���һ��̽��[54]���û��ƿ���ͨ�������ض�miRNA���λ���RNA���յ�����microRNA���⣬�����������ص�³���ԣ�robustness�����м�ֵ�������microRNA���ض�ϸ����֯�б���Ĺ���Ҳ����ת��Ӧ��ǰ����
5.2 ����microRNA�ļ�������
һЩ������Ҫ��microRNA����miR-17~92��miR-155�Լ�miR-122, �Ѿ���֤�������˺ܶ���Ҫ�����ٷ������ܵ��أ������������������߲��ķ�����չ����ˣ��ܶ˾��ʵ��������microRNAΪ�е㣬�о�����microRNA����͵��ص��ֶΡ�miR-122�ڸ�����֯������߱������ά�ָ����л������̬���ڷǾƾ���֬�����ΰ��ȼ����а�����Ҫ��ɫ[55]��Ҳ�����������ٴ����Ƶ�microRNA[56]��Ŀǰ��Ҫ�÷�������ᣨASO��������microRNA���ܣ��������ҩ�↑����չ���������ٴ�ʵ���Ͻ�����ȶ���Ҳ��һЩС���ӻ����ﱻ���ֿ��Ե���microRNA����ˮƽ������ABX464�����ϵ�miR-124�ı���������������ʪ�Թؽ���[57]��
���⣬Disneyʵ���ҿ�����һ������С���ӽ��RNA��ļ������ø�и���߽���RNA�ķ�����RIBOTAC������������microRNAǰ��ı���[58]�����Ǹ÷���������ʵ�����о���ҩ�↑���Σ�ȱ���ٴ�ʵ�����������Ǻ��ֻ���microRNA�������ֶΣ�����Ҫ���ࡢ���Ͻ��������ĵ��о�����ǰ��������microRNA�������õķ��ӻ���Ŀǰ��Ȼû����ȫ���壬���������microRNA����ϸ�����������Ե��о�������Ҫ������Ӧ����ģ�ͣ�չ�����������ͷ���ˮƽ�ϵ�ʵ��[59]��
5.3 ����microRNA�ĺϳ�����ѧ���ػ�·���
microRNA�������еĵ��ط�ʽʹ��߱����л������ˮƽ������һ���ȶ���Χ���������dz����Ϻϳ�����ѧ����Ҫ��֤�ź���������ױ���ˮƽ�����ȶ����ҿ���������ǿ�����˼·����˽�����һЩ�ϳ�����ѧ�ҿ�ʼ��microRNA�������ǵĺϳɻ�·�����[60-61]��������Ҫ���ƵĻ�����ں��Ӳ���microRNA���ڸû����3���DZ����������microRNA�Ľ��λ�㣬�γ�һ��������·�����Ҿ������ϳ��Ժ��죬�������ڶ������ض������ı���ˮƽ��
��microRNA���������Ѿ��д����о��������������Ǵ�������о���Ȼ����δ����������microRNA�ķ��ӻ����о�������Ӧ�÷dz���Ҫ������Ȼ���ص�Զ��microRNA�о�Ҳ��һ�ȺܺõĴ��ڣ�������ѧ���о�mRNA�����ϵĵ��ػ��ƣ�������������ľ����ز��ٽ����ཡ���ͷ�չ������Զ�����塣
���
���ĸ������ߵĻ���ͶԱ����������д�ɡ���Ϊʱ��ִ٣�����δ������������������У���������������ۡ�Ƭ�桢����©֮������������ʵ���ҵ���Ҫ����Ҳδ������һһ���ã��ڴ˱�ʾǸ�⣬����ӭ����ָ����
�����
[1]. Lee, R.C., Feinbaum, R.L., and Ambros, V.��1993��. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 75, 843-854. 10.1016/0092-8674��93��90529-y.
[2]. Wightman, B., Ha, I., and Ruvkun, G.��1993��. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 75, 855-862. 10.1016/0092-8674��93��90530-4.
[3]. Pasquinelli, A.E., Reinhart, B.J., Slack, F., Martindale, M.Q., Kuroda, M.I., Maller, B., Hayward, D.C., Ball, E.E., Degnan, B., Muller, P., et al.��2000��. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature 408, 86-89. 10.1038/35040556.
[4]. Reinhart, B.J., Slack, F.J., Basson, M., Pasquinelli, A.E., Bettinger, J.C., Rougvie, A.E., Horvitz, H.R., and Ruvkun, G.��2000��. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 403, 901-906. 10.1038/35002607.
[5]. Ambros, V.��2008��. The evolution of our thinking about microRNAs. Nat Med 14, 1036-1040. 10.1038/nm1008-1036.
[6]. Ambros, V.��2004��. The functions of animal microRNAs. Nature 431, 350-355. 10.1038/nature02871.
[7]. Lagos-Quintana, M., Rauhut, R., Yalcin, A., Meyer, J., Lendeckel, W., and Tuschl, T.��2002��. Identification of tissue-specific microRNAs from mouse. Curr Biol 12, 735-739. 10.1016/s0960-9822��02��00809-6.
[8]. Calin, G.A., Liu, C.G., Sevignani, C., Ferracin, M., Felli, N., Dumitru, C.D., Shimizu, M., Cimmino, A., Zupo, S., Dono, M., et al.��2004��. MicroRNA profiling reveals distinct signatures in B cell chronic lymphocytic leukemias. Proc Natl Acad Sci U S A 101, 11755-11760. 10.1073/pnas.0404432101.
[9]. Eis, P.S., Tam, W., Sun, L., Chadburn, A., Li, Z., Gomez, M.F., Lund, E., and Dahlberg, J.E.��2005��. Accumulation of miR-155 and BIC RNA in human B cell lymphomas. Proc Natl Acad Sci U S A 102, 3627-3632. 10.1073/pnas.0500613102.
[10]. Lu, J., Getz, G., Miska, E.A., Alvarez-Saavedra, E., Lamb, J., Peck, D., Sweet-Cordero, A., Ebert, B.L., Mak, R.H., Ferrando, A.A., et al.��2005��. MicroRNA expression profiles classify human cancers. Nature 435, 834-838. 10.1038/nature03702.
[11]. Chen, C.Z., Li, L., Lodish, H.F., and Bartel, D.P.��2004��. MicroRNAs modulate hematopoietic lineage differentiation. Science 303, 83-86. 10.1126/science.1091903.
[12]. Tijsterman, M., Ketting, R.F., and Plasterk, R.H.��2002��. The genetics of RNA silencing. Annu Rev Genet 36, 489-519. 10.1146/annurev.genet.36.043002.091619.
[13]. Bernstein, E., Kim, S.Y., Carmell, M.A., Murchison, E.P., Alcorn, H., Li, M.Z., Mills, A.A., Elledge, S.J., Anderson, K.V., and Hannon, G.J.��2003��. Dicer is essential for mouse development. Nat Genet 35, 215-217. 10.1038/ng1253.
[14]. Koralov, S.B., Muljo, S.A., Galler, G.R., Krek, A., Chakraborty, T., Kanellopoulou, C., Jensen, K., Cobb, B.S., Merkenschlager, M., Rajewsky, N., and Rajewsky, K.��2008��. Dicer ablation affects antibody diversity and cell survival in the B lymphocyte lineage. Cell 132, 860-874. 10.1016/j.cell.2008.02.020.
[15]. Muljo, S.A., Ansel, K.M., Kanellopoulou, C., Livingston, D.M., Rao, A., and Rajewsky, K.��2005��. Aberrant T cell differentiation in the absence of Dicer. J Exp Med 202, 261-269. 10.1084/jem.20050678.
[16]. Cobb, B.S., Nesterova, T.B., Thompson, E., Hertweck, A., O'Connor, E., Godwin, J., Wilson, C.B., Brockdorff, N., Fisher, A.G., Smale, S.T., and Merkenschlager, M.��2005��. T cell lineage choice and differentiation in the absence of the RNase III enzyme Dicer. J Exp Med 201, 1367-1373. 10.1084/jem.20050572.
[17]. Ota, A., Tagawa, H., Karnan, S., Tsuzuki, S., Karpas, A., Kira, S., Yoshida, Y., and Seto, M.��2004��. Identification and characterization of a novel gene, C13orf25, as a target for 13q31-q32 amplification in malignant lymphoma. Cancer Res 64, 3087-3095. 10.1158/0008-5472.can-03-3773.
[18]. He, L., Thomson, J.M., Hemann, M.T., Hernando-Monge, E., Mu, D., Goodson, S., Powers, S., Cordon-Cardo, C., Lowe, S.W., Hannon, G.J., and Hammond, S.M.��2005��. A microRNA polycistron as a potential human oncogene. Nature 435, 828-833. 10.1038/nature03552.
[19]. Xiao, C., Srinivasan, L., Calado, D.P., Patterson, H.C., Zhang, B., Wang, J., Henderson, J.M., Kutok, J.L., and Rajewsky, K.��2008��. Lymphoproliferative disease and autoimmunity in mice with increased miR-17-92 expression in lymphocytes. Nat Immunol 9, 405-414. 10.1038/ni1575.
[20]. Ventura, A., Young, A.G., Winslow, M.M., Lintault, L., Meissner, A., Erkeland, S.J., Newman, J., Bronson, R.T., Crowley, D., Stone, J.R., et al.��2008��. Targeted deletion reveals essential and overlapping functions of the miR-17 through 92 family of miRNA clusters. Cell 132, 875-886. 10.1016/j.cell.2008.02.019.
[21]. Rodriguez, A., Vigorito, E., Clare, S., Warren, M.V., Couttet, P., Soond, D.R., van Dongen, S., Grocock, R.J., Das, P.P., Miska, E.A., et al.��2007��. Requirement of bic/microRNA-155 for normal immune function. Science 316, 608-611. 10.1126/science.1139253.
[22]. Thai, T.H., Calado, D.P., Casola, S., Ansel, K.M., Xiao, C., Xue, Y., Murphy, A., Frendewey, D., Valenzuela, D., Kutok, J.L., et al.��2007��. Regulation of the germinal center response by microRNA-155. Science 316, 604-608. 10.1126/science.1141229.
[23]. Vigorito, E., Perks, K.L., Abreu-Goodger, C., Bunting, S., Xiang, Z., Kohlhaas, S., Das, P.P., Miska, E.A., Rodriguez, A., Bradley, A., et al.��2007��. microRNA-155 regulates the generation of immunoglobulin class-switched plasma cells. Immunity 27, 847-859. 10.1016/j.immuni.2007.10.009.
[24]. Dorsett, Y., McBride, K.M., Jankovic, M., Gazumyan, A., Thai, T.H., Robbiani, D.F., Di Virgilio, M., Reina San-Martin, B., Heidkamp, G., Schwickert, T.A., et al.��2008��. MicroRNA-155 suppresses activation-induced cytidine deaminase-mediated Myc-Igh translocation. Immunity 28, 630-638. 10.1016/j.immuni.2008.04.002.
[25]. Monticelli, S., Ansel, K.M., Xiao, C., Socci, N.D., Krichevsky, A.M., Thai, T.H., Rajewsky, N., Marks, D.S., Sander, C., Rajewsky, K., et al.��2005��. MicroRNA profiling of the murine hematopoietic system. Genome Biol 6, R71. 10.1186/gb-2005-6-8-r71.
[26]. Xiao, C., Calado, D.P., Galler, G., Thai, T.H., Patterson, H.C., Wang, J., Rajewsky, N., Bender, T.P., and Rajewsky, K.��2007��. MiR-150 controls B cell differentiation by targeting the transcription factor c-Myb. Cell 131, 146-159. 10.1016/j.cell.2007.07.021.
[27]. Kramer, N.J., Wang, W.L., Reyes, E.Y., Kumar, B., Chen, C.C., Ramakrishna, C., Cantin, E.M., Vonderfecht, S.L., Taganov, K.D., Chau, N., and Boldin, M.P.��2015��. Altered lymphopoiesis and immunodeficiency in miR-142 null mice. Blood 125, 3720-3730. 10.1182/blood-2014-10-603951.
[28]. Boldin, M.P., Taganov, K.D., Rao, D.S., Yang, L., Zhao, J.L., Kalwani, M., Garcia-Flores, Y., Luong, M., Devrekanli, A., Xu, J., et al.��2011��. miR-146a is a significant brake on autoimmunity, myeloproliferation, and cancer in mice. J Exp Med 208, 1189-1201. 10.1084/jem.20101823.
[29]. Kuchen, S., Resch, W., Yamane, A., Kuo, N., Li, Z., Chakraborty, T., Wei, L., Laurence, A., Yasuda, T., Peng, S., et al.��2010��. Regulation of microRNA expression and abundance during lymphopoiesis. Immunity 32, 828-839. 10.1016/j.immuni.2010.05.009.
[30]. Li, Q.J., Chau, J., Ebert, P.J., Sylvester, G., Min, H., Liu, G., Braich, R., Manoharan, M., Soutschek, J., Skare, P., et al.��2007��. miR-181a is an intrinsic modulator of T cell sensitivity and selection. Cell 129, 147-161. 10.1016/j.cell.2007.03.008.
[31]. Mok, Y., Schwierzeck, V., Thomas, D.C., Vigorito, E., Rayner, T.F., Jarvis, L.B., Prosser, H.M., Bradley, A., Withers, D.R., Martensson, I.L., et al.��2013��. MiR-210 is induced by Oct-2, regulates B cells, and inhibits autoantibody production. J Immunol 191, 3037-3048. 10.4049/jimmunol.1301289.
[32]. Li, Y., Deng, X., Zeng, X., and Peng, X.��2016��. The Role of Mir-148a in Cancer. J Cancer 7, 1233-1241. 10.7150/jca.14616.
[33]. Gonzalez-Martin, A., Adams, B.D., Lai, M., Shepherd, J., Salvador-Bernaldez, M., Salvador, J.M., Lu, J., Nemazee, D., and Xiao, C.��2016��. The microRNA miR-148a functions as a critical regulator of B cell tolerance and autoimmunity. Nat Immunol 17, 433-440. 10.1038/ni.3385.
[34]. Bartel, D.P.��2009��. MicroRNAs��target recognition and regulatory functions. Cell 136, 215-233. 10.1016/j.cell.2009.01.002.
[35]. Xiao, C., and Rajewsky, K.��2009��. MicroRNA control in the immune system��basic principles. Cell 136, 26-36. 10.1016/j.cell.2008.12.027.
[36]. Alivernini, S., Gremese, E., McSharry, C., Tolusso, B., Ferraccioli, G., McInnes, I.B., and Kurowska-Stolarska, M.��2017��. MicroRNA-155-at the Critical Interface of Innate and Adaptive Immunity in Arthritis. Front Immunol 8, 1932. 10.3389/fimmu.2017.01932.
[37]. Hsin, J.P., Lu, Y., Loeb, G.B., Leslie, C.S., and Rudensky, A.Y.��2018��. The effect of cellular context on miR-155-mediated gene regulation in four major immune cell types. Nat Immunol 19, 1137-1145. 10.1038/s41590-018-0208-x.
[38]. Kang, S.G., Liu, W.H., Lu, P., Jin, H.Y., Lim, H.W., Shepherd, J., Fremgen, D., Verdin, E., Oldstone, M.B., Qi, H., et al.��2013��. MicroRNAs of the miR-17 approximately 92 family are critical regulators of T��FH��differentiation. Nat Immunol 14, 849-857. 10.1038/ni.2648.
[39]. Jin, H.Y., Oda, H., Lai, M., Skalsky, R.L., Bethel, K., Shepherd, J., Kang, S.G., Liu, W.H., Sabouri-Ghomi, M., Cullen, B.R., et al.��2013��. MicroRNA-17~92 plays a causative role in lymphomagenesis by coordinating multiple oncogenic pathways. EMBO J 32, 2377-2391. 10.1038/emboj.2013.178.
[40]. Lai, M., Gonzalez-Martin, A., Cooper, A.B., Oda, H., Jin, H.Y., Shepherd, J., He, L., Zhu, J., Nemazee, D., and Xiao, C.��2016��. Regulation of B-cell development and tolerance by different members of the miR-17 approximately 92 family microRNAs. Nat Commun 7, 12207. 10.1038/ncomms12207.
[41]. Xie, J., Du, Y., Liu, D., Wu, J., Yang, K., He, X., Zhao, J., Hong, P., Liao, K., Zhang, H., et al.��2023��. The miR-17 approximately 92 miRNAs promote plasma cell differentiation by suppressing SOCS3-mediated NIK degradation. Cell Rep 42, 112968. 10.1016/j.celrep.2023.112968.
[42]. Liao, K., Chen, P., Zhang, M., Wang, J., Hatzihristidis, T., Lin, X., Yang, L., Yao, N., Liu, C., Hong, Y., et al.��2024��. Critical roles of the miR-17 approximately 92 family in thymocyte development, leukemogenesis, and autoimmunity. Cell Rep 43, 114261. 10.1016/j.celrep.2024.114261.
[43]. Jin, H.Y., Oda, H., Chen, P., Yang, C., Zhou, X., Kang, S.G., Valentine, E., Kefauver, J.M., Liao, L., Zhang, Y., et al.��2017��. Differential Sensitivity of Target Genes to Translational Repression by miR-17~92. PLoS Genet 13, e1006623. 10.1371/journal.pgen.1006623.
[44]. Labi, V., Peng, S., Klironomos, F., Munschauer, M., Kastelic, N., Chakraborty, T., Schoeler, K., Derudder, E., Martella, M., Mastrobuoni, G., et al.��2019��. Context-specific regulation of cell survival by a miRNA-controlled BIM rheostat. Genes Dev 33, 1673-1687. 10.1101/gad.330134.119.
[45]. Mukherji, S., Ebert, M.S., Zheng, G.X., Tsang, J.S., Sharp, P.A., and van Oudenaarden, A.��2011��. MicroRNAs can generate thresholds in target gene expression. Nat Genet 43, 854-859. 10.1038/ng.905.
[46]. Chen, P., Liao, K., and Xiao, C.��2018��. MicroRNA says no to mass production. Nat Immunol 19, 1040-1042. 10.1038/s41590-018-0215-y.
[47]. Vasudevan, S., Tong, Y., and Steitz, J.A.��2007��. Switching from repression to activation��microRNAs can up-regulate translation. Science 318, 1931-1934. 10.1126/science.1149460.
[48]. Kato, M.��2023��. Identify MicroRNA Targets Using AGO2-CLASH��Cross-linking, Ligation, and Sequencing of Hybrids��and AGO2-CLIP��Cross-Linking and Immuno-Precipitation��in Cells with or Without the MicroRNA of Interest Depleted. Methods Mol Biol 2666, 137-147. 10.1007/978-1-0716-3191-1_10.
[49]. Griesemer, D., Xue, J.R., Reilly, S.K., Ulirsch, J.C., Kukreja, K., Davis, J.R., Kanai, M., Yang, D.K., Butts, J.C., Guney, M.H., et al.��2021��. Genome-wide functional screen of 3'UTR variants uncovers causal variants for human disease and evolution. Cell 184, 5247-5260 e5219. 10.1016/j.cell.2021.08.025.
[50]. Ameres, S.L., Horwich, M.D., Hung, J.H., Xu, J., Ghildiyal, M., Weng, Z., and Zamore, P.D.��2010��. Target RNA-directed trimming and tailing of small silencing RNAs. Science 328, 1534-1539. 10.1126/science.1187058.
[51]. Cazalla, D., Yario, T., and Steitz, J.A.��2010��. Down-regulation of a host microRNA by a Herpesvirus saimiri noncoding RNA. Science 328, 1563-1566. 10.1126/science.1187197.
[52]. Shi, C.Y., Kingston, E.R., Kleaveland, B., Lin, D.H., Stubna, M.W., and Bartel, D.P.��2020��. The ZSWIM8 ubiquitin ligase mediates target-directed microRNA degradation. Science 370. 10.1126/science.abc9359.
[53]. Han, J., LaVigne, C.A., Jones, B.T., Zhang, H., Gillett, F., and Mendell, J.T.��2020��. A ubiquitin ligase mediates target-directed microRNA decay independently of tailing and trimming. Science 370. 10.1126/science.abc9546.
[54]. Buhagiar, A.F., and Kleaveland, B.��2024��. To kill a microRNA��emerging concepts in target-directed microRNA degradation. Nucleic Acids Res 52, 1558-1574. 10.1093/nar/gkae003.
[55]. Thakral, S., and Ghoshal, K.��2015��. miR-122 is a unique molecule with great potential in diagnosis, prognosis of liver disease, and therapy both as miRNA mimic and antimir. Curr Gene Ther 15, 142-150. 10.2174/1566523214666141224095610.
[56]. Janssen, H.L., Reesink, H.W., Lawitz, E.J., Zeuzem, S., Rodriguez-Torres, M., Patel, K., van der Meer, A.J., Patick, A.K., Chen, A., Zhou, Y., et al.��2013��. Treatment of HCV infection by targeting microRNA. N Engl J Med 368, 1685-1694. 10.1056/NEJMoa1209026.
[57]. Daien, C., Krogulec, M., Gineste, P., Steens, J.M., Desroys du Roure, L., Biguenet, S., Scherrer, D., Santo, J., Ehrlich, H., and Durez, P.��2022��. Safety and efficacy of the miR-124 upregulator ABX464��obefazimod, 50 and 100 mg per day��in patients with active rheumatoid arthritis and inadequate response to methotrexate and/or anti-TNFalpha therapy��a placebo-controlled phase II study. Ann Rheum Dis 81, 1076-1084. 10.1136/annrheumdis-2022-222228.
[58]. Tong, Y., Lee, Y., Liu, X., Childs-Disney, J.L., Suresh, B.M., Benhamou, R.I., Yang, C., Li, W., Costales, M.G., Haniff, H.S., et al.��2023��. Programming inactive RNA-binding small molecules into bioactive degraders. Nature 618, 169-179. 10.1038/s41586-023-06091-8.
[59]. Kilikevicius, A., Meister, G., and Corey, D.R.��2022��. Reexamining assumptions about miRNA-guided gene silencing. Nucleic Acids Res 50, 617-634. 10.1093/nar/gkab1256.
[60]. Strovas, T.J., Rosenberg, A.B., Kuypers, B.E., Muscat, R.A., and Seelig, G.��2014��. MicroRNA-based single-gene circuits buffer protein synthesis rates against perturbations. ACS Synth Biol 3, 324-331. 10.1021/sb4001867.
[61]. Du, R., Flynn, M.J., Honsa, M., Jungmann, R., and Elowitz, M.B.��2024��. miRNA circuit modules for precise, tunable control of gene expression. bioRxiv. 10.1101/2024.03.12.583048.