针对付向东教授提出的几点质疑,我的回复如下:
1. 付向东教授在神经所做的报告是采用小分子抑制剂的方法,而他们在Nature发表的论文采用的却是shRNA和ASO的方法。连他自己在Nature文章中都没有提及他在报告中采用的小分子抑制剂的方法。他又是如何在当时向我们透入大量的技术细节的?他报告中的方法最终证明是错误或不可行的,难道已经公开发表5年的位点就可以霸占,不允许其他人用新的技术来尝试吗?这和大佬圈地有何区别呢?
2. 根据付教授发表的文章和公布的专利(2018年4月递交,2019年10月公开),丝毫没有提及用基因编辑方法治疗帕金森病。而且他还宣称在我们注射的纹状体脑区通过在星形胶质细胞内敲低Ptbp1几乎不可能形成多巴胺神经元。而我们恰恰是在纹状体中高效诱导出多巴胺神经元产生,进而达到治疗效果。在举报信中,他通过混淆视听,让大家认为我完全抄袭他的结果。
3. 付教授提到,他还分享了Ptbp1应用到视网膜疾病治疗的工作,首先我确实不知道他的分享,其次,最近我们通过BioRxiv检索,也找到2020年4月8日在线刊登的这篇文章。他们从实验方法到动物疾病模型没有一处一样,更重要的是他们的目的是将穆勒胶质细胞转分化为视锥细胞(Cone),而不是我们文章中的目的神经元视神经节细胞。两篇文章的连实验目的都不一样,何来剽窃?付向东教授专利中涵盖的疾病和方法,并没有涵盖任何RGC细胞的转分化。可见他是如何向我透入这些技术细节。最后我想指出的是,我们Cell文章主要关注点是视神经节细胞的转分化(7个主图中有5个与视神经节细胞的转分化有关)。
4. 关于我造假的言论,纯属污蔑。我2013年Cell文章已经有许多实验室重复出我们的结果和方法,详见下文。他为了抹黑我,连基本的科学事实都不顾,这不是一个正派的科学家做的事情。
关于本人2020年Cell文章抄袭和剽窃美国加利福尼亚大学圣迭戈分校付向东教授文章、造假的争论1. 付向东教授在神经所报告中提到的Ptbp1靶点,已经于2013年发表。他报告中提到利用小分子抑制剂靶向Ptbp1进行体内转分化的相关研究。但是我们认为小分子药物既没有强大的分子靶向特异性,又没有足够的细胞靶向特异性,因此我们也意识到我们擅长的基因编辑策略具有很多的优势。RNA靶向基因编辑技术不仅可以实现细胞靶向特异性,而且在体内可以实现高效特异性编辑。难道已经公开发表5年的位点就不允许其他人用新的技术来尝试吗?而且在他公布的专利中并没有涵盖利用基因编辑技术来实现转分化,可见他们团队当时并不是十分了解基因编辑技术。2. 付向东教授说向我透漏了许多实验细节。我们希望他能提供任何支持他观点的证据。事实上,付向东教授在神经所做的报告是采用小分子抑制剂的方法,而他们在Nature发表的论文采用的却是shRNA和ASO的方法。连他自己在Nature文章中都没有提及他在报告中采用的小分子抑制剂的方法。他们又是如何在当时向我们透入大量的技术细节的?他报告中的结果最终证明是错误或不可行的,难道能让所有其他人都不能尝试他已经2013年报道的靶点吗?这和大佬圈地有何区别呢?其次,根据付教授发表的文章和在我们文章投稿后公布的专利(公布时间:2019年10月17日),他发现在我们注射的纹状体脑区几乎不可能形成多巴胺神经元。而我们恰恰是在纹状体中高效诱导出多巴胺神经元产生,进而达到治疗效果。我们实验室从2014年至今一直致力于基因编辑技术在各个领域的应用以及安全性评价。这项工作的主要完成人为我实验室博士后周海波,他在获得神经科学博士学位后来到我实验室,在神经研究方面具有非常强的背景和训练。我们15年开始优化并测试利用基因编辑技术进行体内星形胶质细胞向神经元转分化,并于2018年一月份将此项工作发表在Nature Neuroscience上。由于我们这篇文章中使用的基因激活元件太大,很难实现体内运输,导致很难应用此技术进行神经元转分化通过成体治疗的方式治疗神经系统疾病。但我们一直都没有就此停止探索其他利用基因编辑技术方式进行神经元转分化的策略。2013年付向东教授在Cell上发表的文章证明,通过shRNA敲低Ptbp1基因就可以实现各种细胞向神经元的转分化。虽然还有少数其他通过被抑制就可以实现神经元转分化的靶点,但它们效率都很低,这点也在付向东教授的文章中进行了比较。因此, Ptbp1可以说是目前唯一个通过敲低就可以高效实现神经元转分化的靶点,在领域内非常著名。虽然考虑通过基因编辑抑制基因表达来实现神经元转分化,但由于当时没有合适的基因编辑工具出现,该想法就暂时被搁置。2018年3月15日,CasRx在Cell上发表,显示极其高效的mRNA敲低效率,并且相对于shRNA技术具有更强的特异性。同时,CasRx体系足够小,非常适合通过AAV递送到体内进行靶向治疗。这项技术与以往的shRNA相比展现了极大的治疗潜力,我们认为小分子药物既没有强大的分子靶向特异性,又没有足够细胞的靶向特异性,因此我们敏感的察觉到我们擅长的基因编辑策略具有很多的优势。所以我们迅速跟进。我们首先针对外源性的荧光蛋白进行了测试,并于2018年5月初得到了非常好的敲低结果。其后我们又针对一个内源性的治疗靶点Vegfa进行了测试,因为该靶点是被验证过的年龄相关性黄斑变性的治疗靶点,而眼睛又是我们非常擅长递送AAV且容易检测的系统。2018年5月,我们首先在最擅长且最容易运输的眼睛系统中检测CasRx是否能在体内实现高效的mRNA敲低。2018年7月初我们获得了非常好的CasRx体内敲低数据,之后我们全面铺开在眼睛、耳朵、肝脏和大脑中对包括Ptbp1在内的已经明确发表的治疗靶点进行大量测试。(2018年5月7号,我们首次在实验室证明CasRx可以在细胞内靶向过表达的荧光蛋白mCherry,详细数据请见附件一。随后我们开始针对能够通过敲低实现疾病治疗的多个基因靶点进行了测试。)2018年5月25日,我们拿到了通过CasRx在293T细胞内能够高效的敲低黄斑病变的治疗靶点VEGFA的数据。2018年5月31日,我们拿到细胞内的数据,证明CasRx可以在对视网膜色素变性治疗靶点Rho的点突变的RNA进行敲低。2018年6月9日,在细胞内拿到CasRx能够对遗传性耳聋的治疗靶点Tmc1敲低的数据。2018年6月20日,在细胞内证明CasRx能够对天使综合征的治疗靶点Ube3a-ATS进行敲低。2018年6月27日,在细胞内证明CasRx能够对脊髓性肌萎缩症(SMA)的治疗靶点SMN_AS1进行敲低。2018年7月12日,在N2a细胞内证明CasRx能够对帕金森综合征的治疗靶点Ptbp1进行敲低。2018年9月15日,在小鼠的N2A细胞内证明CasRx能够靶向黄斑病变另外一个治疗靶点Hif1a。2018年11月3日,设计了CasRx靶向一型糖尿病的治疗靶点HPD。Reviewer comments:“In particular, previous efforts of RNA-targeting CRISPR systems focused on knockdown of toxic mutant transcripts in models of mendelian inherited genetic disease, whereas this study presents the novel aspect of using these tools for in vivo therapeutic cell fate conversion, which is quite interesting and could be a broadly applicable approach.”译文:审稿人评论:“尤其是,以往的研究都集中在利用RNA靶向CRISPR系统来直接降低遗传病模型中有害的突变转录本,而这项研究却利用这些工具在体内进行治疗性的细胞命运转分化,从而展现一个全新的视角,这是非常有意思的,而且可以得到广泛的应用。” Reviewer comments: The concept of converting already present precursor cells to neurons that are capable of integrating and extending axons is extremely valuable。“把已经存在的前体细胞直接转化为拥有可以整合和延伸的轴突的神经元,这个概念极其的有价值。”付向东教授于2018年6月14日来到神经所做报告。我们了解到,付向东教授在利用小分子抑制剂靶向Ptbp1进行体内转分化的相关研究。我们一直认为小分子药物既没有强大的分子靶向特异性,又没有足够细胞的靶向特异性,因此我们仍一直坚持我们自己原有的基因编辑策略进行体内转分化研究。后来我们从付向东教授后来的bioRvix中得知,他们用的方法是shRNA和ASO。关于抄袭的诬告,付教授声称把所有技术细节都告诉我了,希望他能提供任何支持他观点的证据。根据付教授发表的文章和公布的专利(在我们投稿后公布),他发现在我们注射的纹状体脑区几乎不可能形成多巴胺神经元,而我们的结论与之恰恰相反,我们是在纹状体中高效诱导出多巴胺神经元产生,进而达到治疗效果。需要指出的是付向东教授的文章通过在GFAP-Cre转基因小鼠结合Cre-dependent的shRNA实现帕金森症状的治疗,但是此项技术在临床上是无法应用的。付向东教授还使用了ASO策略,但这种方法无法实现细胞靶向特异性。我们的文章,通过RNA靶向基因编辑技术不仅可以实现细胞靶向特异性,而且在帕金森小鼠模型上实现了非常好的治疗效果。同时,我们的基因编辑方法可以在不同的非基因修饰疾病动物模型中进行应用,展示了强大的临床潜力。关于穆勒胶质细胞标记的特异性低导致的误标,我们的数据已经显示可以实现非常特异的细胞靶向性。而且我们也非常乐意分享我们的方法以及材料,希望全世界所有感兴趣的科学家进行重复。
付教授提到,他还分享了Ptbp1应用到视网膜疾病治疗的工作,我们通过BioRxiv检索,也找到2020年4月8日在线刊登的,美国加州圣地亚哥大学的Kang Zhang与付向东教授合作的文章Visual function restoration in genetically blind mice via endogenous cellular reprogramming。

该文章中用到的方法、小鼠模型、质粒、病毒都与我们不同。而且,他们的研究是Rd10小鼠 (一种视锥细胞和视杆细胞退化的小鼠) 中进行的。最重要的是,他们的目的是将Muller细胞转分化为视锥细胞(Cone),而不是视网膜神经节细胞。两篇文章的连目的都不一样,更没有一样的数据。关于因GFAP-Cre可能会在RGC中表达泄露而怀疑我们做假,我认为,生物实验最重要的是尊重实验结果,而不是根据猜测或者 “众所周知”而下结论。面对这个学术争议,我立刻让实验室其他学生重复了这个实验,实验结果很好的验证了我们结论的可靠性。这一实验并不复杂,转基因小鼠是常用的类型,国内外许多实验室都可及;注射的病毒商业化的公司都可以提供,可现货购买。小鼠和病毒准备好,只需要2-3周的时间就能验证我们的结果是否可靠。我们文章已发表2个多月,我们欢迎所有实验室重复我们的实验结果并讨论。更有意思的是,付向东教授和Kang Zhang合作的文章中也用到GFAP-Cre的AAV病毒系统进行标记,并声明能用GFAP-Cre特异性标记胶质穆勒细胞(见下图)。而且需要指出的是,他们用的是GFAP-Cre+CMV-LSL-RFP 双AAV病毒系统,我们用的是GFAP-Cre单AAV病毒系统注射到Ai9小鼠中。如果我也来一个“众所周知”,双病毒系统比单病毒系统更容易泄露,那我也有理由怀疑这篇文章中的结果存在造假的可能。我是否也可以怀疑他们要么观察到了泄露现象而不实报道,要么没有观察到泄露现象而故意抹黑我们,希望他们能够正面回应一下。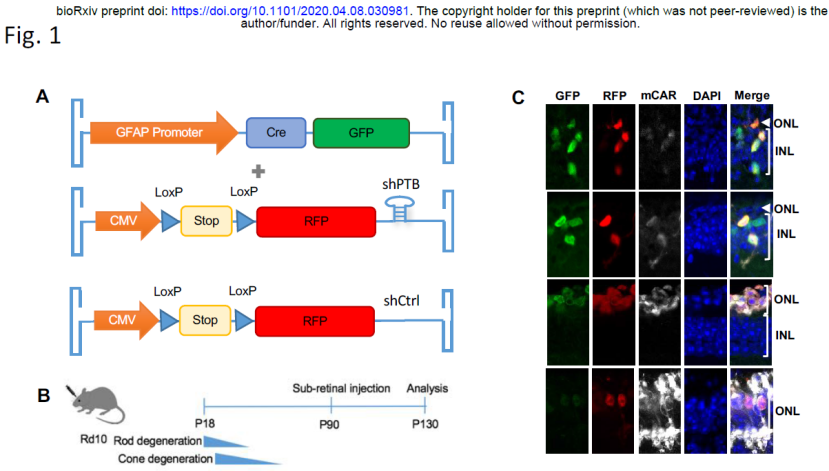 付向东教授Nature文章专利申请号WO2019200129A1,公布时间:2019年10月17日(在我们Cell文章投稿后公布)https://patents.google.com/patent/WO2019200129A1/en?inventor=xiang-dong+fu
付向东教授Nature文章专利申请号WO2019200129A1,公布时间:2019年10月17日(在我们Cell文章投稿后公布)https://patents.google.com/patent/WO2019200129A1/en?inventor=xiang-dong+fu 付向东教授Nature文章和专利中都指明在纹状体(striatum)中几乎不可能实现多巴胺神经元转分化:
付向东教授Nature文章和专利中都指明在纹状体(striatum)中几乎不可能实现多巴胺神经元转分化:

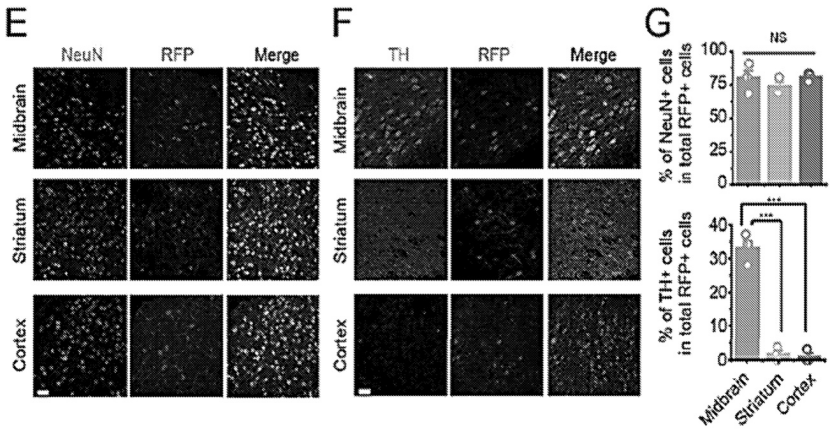
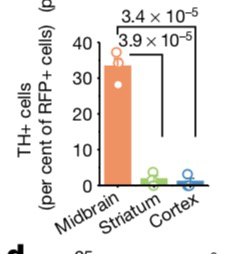 付向东教授专利中涵盖的疾病和方法,并没有涵盖视神经节细胞转分化和基因编辑技术:
付向东教授专利中涵盖的疾病和方法,并没有涵盖视神经节细胞转分化和基因编辑技术:

关于本人2013年Cell文章造假部分回应
“这让我联想到杨辉在博士后阶段发表的“高效插入基因突变方法”研究论文 (详见 Cell 154:1370-1379, 2013),遭到领域内科学家广泛质疑,有20多个独立实验室联合报道不能重复他的实验结果 (详见 Genome Biology 20:171, 2019) 。对于其他科学家的质疑,杨辉除了辩称自己比别人高明,强调实验条件略有不同外,没有任何合理解释,至今我们再也没有看到他重复自己结果的实验证据。”Gurumurthy et al.等在2019年发表的文章中称我们在2013年Cell文章中制作条件性敲除小鼠的方法效率比文中报道的要低很多,而且比较了几种制作条件性敲除的新方法。这是很正常的学术争论,同时我们并不同意文中的一些观点,也写信给杂志社回应了作者,文章正在审稿中,由于疫情原因耽误了发表,原文如下:Response to “Reproducibility of CRISPR-Cas9 methods for generation of conditional mouse alleles:a multi-center evaluation”Hui Yang【1】, Haoyi Wang【2】, Rudolf Jaenisch【3,4】【1】Institute of Neuroscience, State Key Laboratory of Neuroscience, Key Laboratory of Primate Neurobiology, CAS Center for Excellence in Brain Science and Intelligence Technology, Shanghai Research Center for Brain Science and Brain-Inspired Intelligence, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, Shanghai 200031, China【2】State Key Laboratory of Stem Cell and Reproductive Biology, Institute of Zoology, The Chinese Academy of Sciences, Beijing, China【3】Whitehead Institute for Biomedical Research, Cambridge, MA, USA 4Department of Biology, Massachusetts Institute of Technology, Cambridge, MA, USA.*Corresponding authors. E-mail:huiyang@ion.ac.cn; wanghaoyi@ioz.ac.cn; jaenisch@wi.mit.edu
Gurumurthy et al. [1] recently reported that a method developed by Yang et al. to generate floxed allele(designated as “two donor method” by Gurumurthy et al.)[2] had poor reproducibility. They claimed that three centers could not reproduce our results on generating conditional alleles of the Mecp2 locus and that the “two-donor method” had very low success rate on other loci.Here, we provide our responses to these claims:1. Our results on Mecp2 locus published by Yang et al have been reproduced by independent experiments in the Jaenisch(8-10% correct alleles), Yang(8% correct alleles)and Hatada’s groups(2-6% correct alleles)[3], respectively. In addition, multiple peer-reviewed publications [3, 9-12] have successfully used this method to create conditional knockout(CKO)mice(9 out of 11 loci succeeded, 2.5% to 18% efficiency). We noticed that the efficiency of generating CKO mice by CRISPR/Cas9 could vary, which might due to different platform features or experiment conditions.2. The conditions used by Gurumurthy et al. [1] do not correspond to the conditions used in our paper. The concentrations of CRISPR reagents used in the Gurumurthy et al.’s study [1] on the Mecp2 locus(10 ng/μl for Cas9 mRNA, 10 ng/μl for sgRNA and 10 ng/μl for oligos)were much lower(10 fold lower RNA and 20 fold lower oligo donor concentration)than those used in the Yang et al.’s experiments(Cas9 100 ng/μl, sgRNA 50 ng/μl and 100 ng/μl for each oligo)[2] and Yang et al.’s previous [4] and following publications [5-8]. It is well known that the concentrations of CRISPR reagents are well correlated with the genome editing efficiency.3. We utilized piezo-driven zygote injection method in our original paper, which allows for injecting CRISPR components at much higher concentration. The difference between this method and pronuclear injection method used by Gurumurthy et al. might also contribute to the difference of successful rates.In general, with any genome editing method or strategy being used, the efficiencies at different genomic loci are often highly variable. In the 2013 proof of concept paper, we showed the feasibility of generating floxed allele at Mecp2 locus using CRISPR. To assume the efficiency we demonstrated at Mecp2 locus will be directly translated to the success rate at other genomic loci seems premature.We agree with the Gurumurthy et al’s comment that the “one-donor method” offers higher success rate for generating floxed alleles in general, while the efficiency of “one-donor method” is also variable depending on the genomic loci and donor plasmid design. Before the publication of Gurumurthy et al., we also noted this, and developed a “one-donor method”, termed “Tild-CRISPR” method [8], and demonstrated the feasibility and high efficiency in generating CKO mice.With the fast improvement of genome editing technologies, we and many others constantly optimize our protocols. We welcome all discussions about the choice of optimal strategy for particular applications, however, we think the reproducibility of any published work can only be validated by using the exact same experimental methods and technical parameters.Gurumurthy等最近报道说由Yang开发的用于产生floxed等位基因的方法重现性较差(Gurumurthy指定该方法为“双供体法”) [1-2]。他们声称有三个实验中心无法重复在Mecp2位点产生条件等位基因的结果,同时“双供体法”在其他位点的成功率非常低。在这里,我们对Gurumurthy等的说法解释如下:1.我们在Mecp2位点的结果已经被三个不同的实验室分别重复出来:Jaenisch实验室(8-10%正确等位基因)、Yang实验室(8%正确等位基因)和Hatada实验室(2-6%正确等位基因)[3]。此外,多篇同行评议论文已经成功地使用该方法创建了条件敲除(CKO)小鼠(11个基因中有9个成功,效率为2.5% - 18%不等)[3,9 -12]。我们认为CRISPR/Cas9生成CKO小鼠的效率之所以存在差异,这与平台技术或实验条件的不同有关。2.众所周知,基因组编辑效率与CRISPR试剂的浓度密切相关。而Gurumurthy等使用的CRISPR试剂的浓度条件与我们文章中使用的条件不一致[1],Gurumurthy等使用的注射材料的浓度比我们要低得多(10-20倍差异) [5 -8],具体如下表所示。3.在我们2013年的论文中,我们使用piezo驱动的受精卵注射法,该方法允许进行高浓度的CRISPR组份注射。而该方法与Gurumurthy等人使用的原核注射方法不同,这可能也是成功率不同的原因。一般来说,任何基因组编辑方法或策略在不同基因组位点的效率往往是高度不同的。在2013年的文章中,我们展示了利用CRISPR在Mecp2位点生成floxed等位基因的可行性,但并不能直接转化为在其他基因组位点上也可以获得相似的成功率。我们同意Gurumurthy等的观点,即一般情况下“单供体方法”在产生floxed等位基因方面展示了更高的成功率,但“单供体方法”的效率也取决于基因组位点和供体质粒设计等因素。在Gurumurthy等的工作发表之前,我们也注意到了这一点并开发了一种名为Tild-CRISPR的“单供体方法” [8],并证明此方法可以高效方便地生成CKO小鼠。随着基因编辑技术的快速发展,我们和其他同行在不断优化我们的策略。我们欢迎所有关于特定情况下选择最佳策略的讨论。但我们认为,任何已发表工作的再现性只能通过使用完全相同的实验方法和技术参数来进行验证。评判生物实验结果真实性很重要的一点是可重复性。既然有许多实验室已经通过我们的方法重复出相同的结果,就不能定义我们的文章是造假。生物实验的复杂性使得任何一个关键因素的差别都可能会极大影响实验结果。如果一些实验室因为各种原因无法重复我们的实验,就忽视重复出我们实验的课题组,并断定文章造假,那么任何一个引用率高的文章都可能会被冠以造假的头衔。
1.Gurumurthy CB, O'Brien AR, Quadros RM, Adams J, Jr., Alcaide P, Ayabe S, Ballard J, Batra SK, Beauchamp MC, Becker KA, et al:Reproducibility of CRISPR-Cas9 methods for generation of conditional mouse alleles:a multi-center evaluation. Genome Biol 2019, 20:171.
2.Yang H, Wang H, Shivalila CS, Cheng AW, Shi L, Jaenisch R:One-Step Generation of Mice Carrying Reporter and Conditional Alleles by CRISPR/Cas-Mediated Genome Engineering. Cell 2013, 154:1370-1379.
3.Horii T, Morita S, Kimura M, Terawaki N, Shibutani M, Hatada I:Efficient generation of conditional knockout mice via sequential introduction of lox sites. Sci Rep 2017, 7:7891.
4.Wang H, Yang H, Shivalila CS, Dawlaty MM, Cheng AW, Zhang F, Jaenisch R:One-step generation of mice carrying mutations in multiple genes by CRISPR/Cas-mediated genome engineering. Cell 2013, 153:910-918.
5.Yang H, Wang H, Jaenisch R:Generating genetically modified mice using CRISPR/Cas-mediated genome engineering. Nat Protoc 2014, 9:1956-1968.
6.Yao X, Wang X, Hu X, Liu Z, Liu J, Zhou H, Shen X, Wei Y, Huang Z, Ying W, et al:Homology-mediated end joining-based targeted integration using CRISPR/Cas9. Cell Res 2017, 27:801-814.
7.Yao X, Liu Z, Wang X, Wang Y, Nie YH, Lai L, Sun R, Shi L, Sun Q, Yang H:Generation of knock-in cynomolgus monkey via CRISPR/Cas9 editing. Cell Res 2018, 28:379-382.
8.Yao X, Zhang M, Wang X, Ying W, Hu X, Dai P, Meng F, Shi L, Sun Y, Yao N, et al:Tild-CRISPR Allows for Efficient and Precise Gene Knockin in Mouse and Human Cells. Dev Cell 2018, 45:526-536 e525.
9.Pritchard CEJ, Kroese LJ, Huijbers IJ:Direct Generation of Conditional Alleles Using CRISPR/Cas9 in Mouse Zygotes. Methods Mol Biol 2017, 1642:21-35.
10.Bishop KA, Harrington A, Kouranova E, Weinstein EJ, Rosen CJ, Cui X, Liaw L:CRISPR/Cas9-Mediated Insertion of loxP Sites in the Mouse Dock7 Gene Provides an Effective Alternative to Use of Targeted Embryonic Stem Cells. G3(Bethesda)2016, 6:2051-2061.
11.Ma X, Chen C, Veevers J, Zhou X, Ross RS, Feng W, Chen J:CRISPR/Cas9-mediated gene manipulation to create single-amino-acid-substituted and floxed mice with a cloning-free method. Sci Rep 2017, 7:42244.
12.Nakagawa Y, Sakuma T, Nishimichi N, Yokosaki Y, Yanaka N, Takeo T, Nakagata N, Yamamoto T:Ultra-superovulation for the CRISPR-Cas9-mediated production of gene-knockout, single-amino-acid-substituted, and floxed mice. Biol Open 2016, 5:1142-1148.