ЩјЭИаВЦШЃЌШчИЩКЕКЭИпбЮЖШЃЌЖджВЮяЩњГЄКЭЩњВњСІВњЩњВЛРћгАЯьЁЃЭбТфЫсЃЈABAЃЉдкЩјЭИаВЦШЯТЛ§РлЃЌЭЈЙ§ABAаХКХДЋЕМЭЈТЗв§ЗЂЖржжЩњРэЗДгІЃЌдіЧПжВЮяЕФПЙФцадЁЃIIIРр SNF1ЯрЙиЕААзМЄУИ 2 ЃЈSnRK2sЃЉ ЪЧABAаХКХДЋЕМЕФЙиМќЕїПивђзгЁЃОЁЙмSnRK2sГЄЦквдРДБЛШЯЮЊЪЧдк PP2CНщЕМЕФвжжЦжаЪЭЗХКѓЭЈЙ§здСзЫсЛЏЖјВњЩњздМЄЛюЃЌЕЋзюНќЗЂЯжЫќУЧдкЩјЭИаВЦШЬѕМўЯТБЛBРрRafбљМЄУИЕФСНИіЖРСЂбЧМвзхЃЌB2 RAFsКЭB3 RAFsМЄЛюЁЃШЛЖјЃЌетаЉRAFЖдSnRK2ЕФСзЫсЛЏКЭSnRK2здЩэСзЫсЛЏжЎМфЕФЙиЯЕШдШЛЪЧЮДжЊЕФЁЃ
2023Фъ6дТ5ШеЃЌPNASдгжОдкЯпЗЂБэСЫШеБОЖЋОЉДѓбЇЕФвЛЦЊЬтЮЊЁАConstitutively active B2 Raf-like kinases are required for drought-responsive gene expression upstream of ABA-activated SnRK2 kinases ЁБЕФбаОПТлЮФЁЃИУбаОПБэУїЃЌЕБДгPP2CНщЕМЕФвжжЦжаЪЭЗХЪБЃЌB2-RAF RAF11ГжајЛюдОВЂМЄЛюSnRK2sЃЌЖјB3-RAF RAF5дкгІМЄЬѕМўЯТБЛМЄЛюЁЃвђДЫЃЌSnRK2здЩэСзЫсЛЏВЛзувдВњЩњABAЗДгІЃЌВЂЧвB2 RAFsЪЧЧсЖШИЩКЕЬѕМўЯТSnRK2МЄЛювдЯьгІABAЕФБивЊЬѕМўЃЌЖјB3 RAFsЪЧбЯжиЩјЭИаВЦШЯТSnRK2ЛюадЕФдіЧПМСЁЃ

БОбаОПЗЂЯжRAF7ЁЂRAF10ЁЂRAF11КЭRAF12етЫФжжМЄУИЖМЪєгк RAFбљЕААзМЄУИЕФB2бЧМвзхЁЃгаШЄЕФЪЧетЫФжжB2 - RAFЖМАќКЌдкетаЉКђбЁЕФIIIбЧРрSnRK2ЯрЛЅзїгУЕААзжаЃЌЕЋВЛАќРЈB3 - RAFбљМЄУИЁЃетБэУїB2КЭB3 - RAFФмдкЩјЭИаВЦШЬѕМўЯТвдВЛЭЌЕФЗНЪНЕїПиIIIРрSnRK2sЁЃ
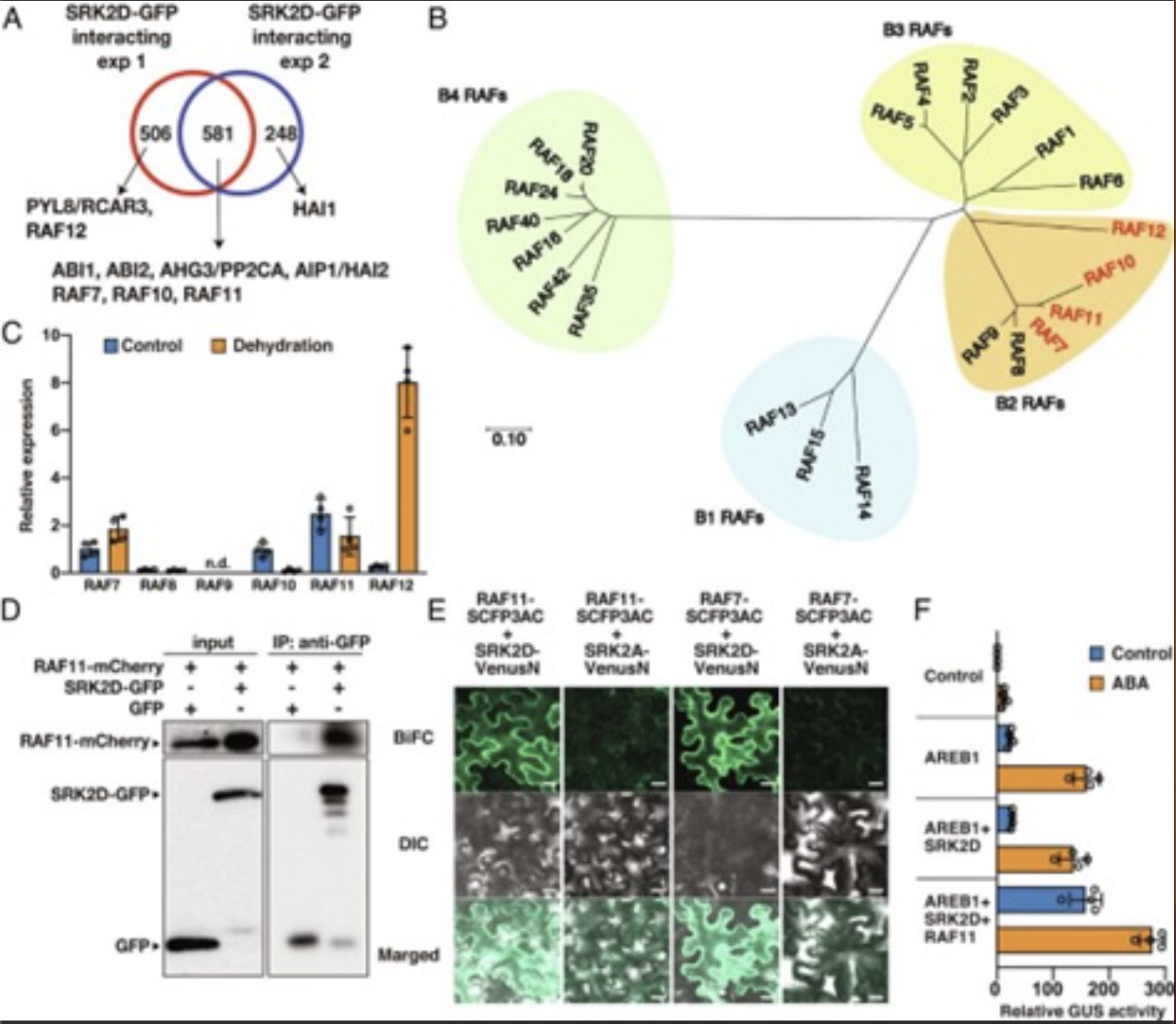
ЭМ1ЃК4ИіB2 RAFБЛМјЖЈЮЊIIIбЧРрSnRK2ЯрЛЅзїгУЕААз
дкИЩКЕаВЦШЯТЃЌЗЂЯжЫФжжB2-RAF, RAF7ЁЂRAF10ЁЂRAF11КЭRAF12ЪЧSnRK2sвРРЕABAМЄЛюЫљБиашЕФЃЌЬхЭтСзЫсЛЏЪЕбщБэУїЫќУЧжБНгМЄЛюIIIРрSnRK2sЃЈЭМ2 A-CЃЉЃЌгывАЩњаЭЯрБШЃЌЭбЫЎКЭABAДІРэЬѕМўЯТЃЌraf7/10/11/12ЫФжиЭЛБфЬхжаIIIРрSnRK2sЕФЛюадЯджјНЕЕЭЃЈЭМ2 F-HЃЉЁЃетаЉЗЂЯжБэУїЃЌIIIРрSnRK2sЕФздСзЫсЛЏВЛзувдВњЩњABAЗДгІЃЌетаЉB2-RAFашвЊвдABAвРРЕЕФЗНЪНМЄЛюIIIРрSnRK2sЁЃСзЫсЛЏЕФНсЙћЗжЮіБэУї,B3-RAF RAF5ЭбЫЎаВЦШВХМЄЛю,ЖјB2-RAF RAF11ГжајЛюдОВЛЙмбЙСІЬѕМўЛђABAЕФДцдкЃЈЭМ2 DКЭEЃЉЁЃ
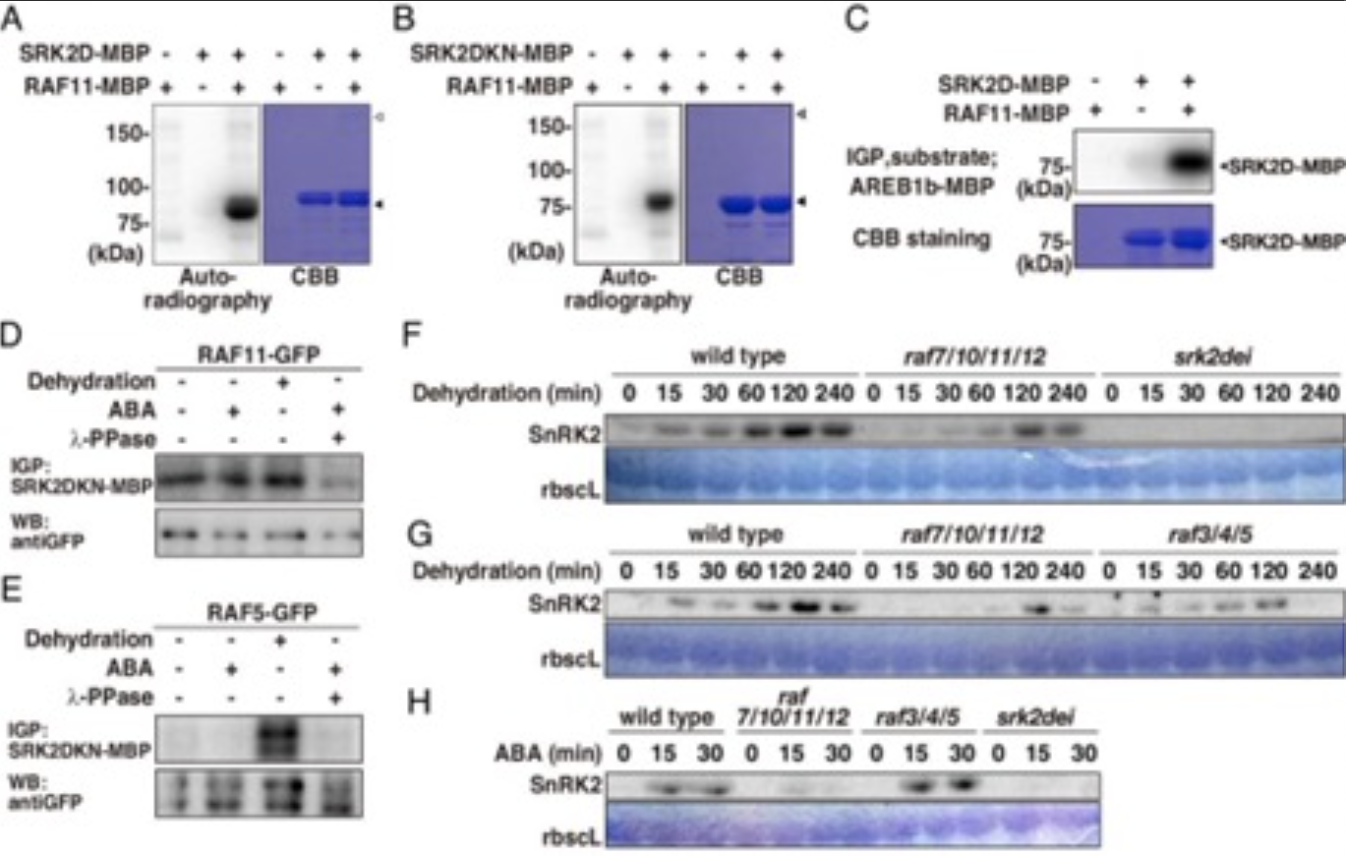
ЭМ2ЃКЫФИіB2 RAFМЄЛюIII РрSnRK2sвдЯьгІЭбЫЎКЭABA
ЮЊСЫВћУїB2-RAFsдкжВЮяИЩКЕКЭABAЯьгІжаЕФЩњРэзїгУЃЌБШНЯСЫвАЩњаЭЁЂraf7/10/11/12ЁЂsrk2deiКЭABAШБЗІaba2-1ЭЛБфЬхжВЮяЕФПЙКЕадЁЃОЁЙмШБЗІB2-RAFЕФЭЛБфЬхraf7/10/11/12БэЯжГіУїЯдЕФABAВЛУєИаБэаЭЃЌЕЋШБЗІB3-RAFЕФЭЛБфЬхraf3/4/5НіБэЯжГіШѕЕФABAВЛУєИаБэаЭЃЈЭМ3 D-FЃЉЁЃетаЉНсЙћБэУїЃЌЮоТлгІМЄЬѕМўШчКЮЃЌB2-RAFЖМЪЧЛюдОЕФЃЌB2-RAFЛсСЂМДСзЫсЛЏВЂМЄЛюIIIРрSnRK2sЁЃЯрБШжЎЯТЃЌB3-RAFдкЭбЫЎаВЦШЯТБЛЧПСвМЄЛюЃЌВЂвдВЛвРРЕABAЕФЗНЪНдіЧПIIIРрSnRK2sЕФЛюадЁЃвђДЫЃЌB2-RAFsКЭB3-RAFsетСНИібЧМвзхдкABAаХКХДЋЕМжаЕФзїгУЪЧВЛЭЌЕФЁЃ
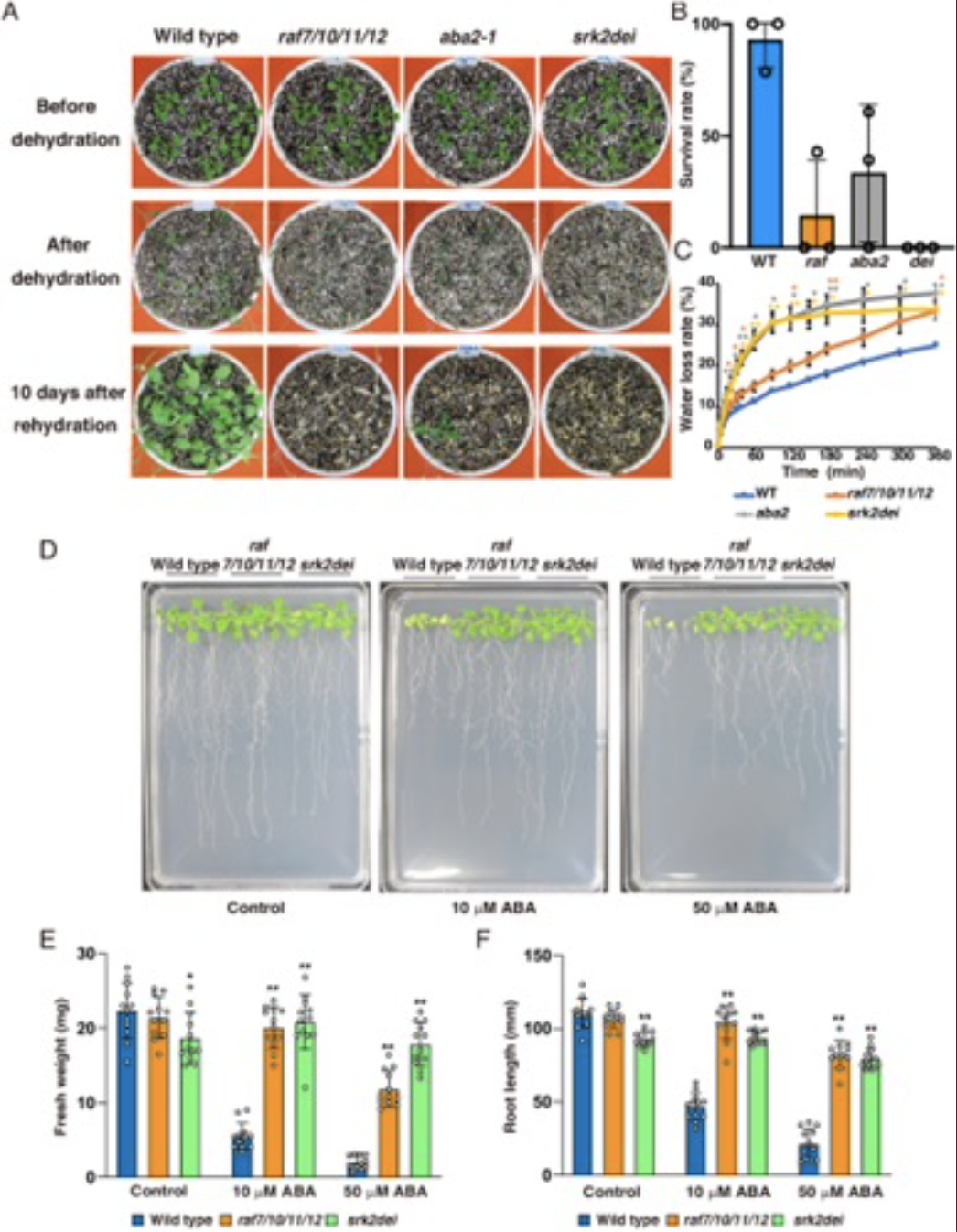
ЭМ3ЃКraf7/10/11/12дкИЩКЕаВЦШЛђИпABAЬѕМўЯТЕФЩњГЄБэаЭ
ИУбаОПЬсГіСЫИЩКЕаВЦШЬѕМўЯТЕФABAаХКХФЃаЭЃЌЦфжаIIIРрSnRK2sЯьгІABAЕФздСзЫсЛЏВЛЙЛГфЗжЃЌашвЊЩЯгЮМЄУИRAF7ЁЂRAF10ЁЂRAF11КЭRAF12ВХФмМЄЛюЃЌIIIРрSnRK2sБЛГжајЛюадзДЬЌЕФB2-RAFСзЫсЛЏЃЌЩѕжСдкЗЧгІМЄЯИАћжавВБЛМЄЛюЁЃЕБИЩКЕаВЦШБфЕУИќбЯжиЪБЃЌжВЮяжаЕФаэЖрЯИАћНгЪеЕНаВЦШаХКХЃЌB3-RAFБЛМЄЛюЃЌВЂжБНгМЄЛюIIIбЧРрSnRK2sЃЌДгЖјдіЧПаВЦШЯьгІЁЃ

ЭМ3ЃКraf7/10/11/12дкИЩКЕаВЦШЛђИпABAЬѕМўЯТЕФЩњГЄБэаЭ
ИУбаОПЬсГіСЫИЩКЕаВЦШЬѕМўЯТЕФABAаХКХФЃаЭЃЌЦфжаIIIРрSnRK2sЯьгІABAЕФздСзЫсЛЏВЛЙЛГфЗжЃЌашвЊЩЯгЮМЄУИRAF7ЁЂRAF10ЁЂRAF11КЭRAF12ВХФмМЄЛюЃЌIIIРрSnRK2sБЛГжајЛюадзДЬЌЕФB2-RAFСзЫсЛЏЃЌЩѕжСдкЗЧгІМЄЯИАћжавВБЛМЄЛюЁЃЕБИЩКЕаВЦШБфЕУИќбЯжиЪБЃЌжВЮяжаЕФаэЖрЯИАћНгЪеЕНаВЦШаХКХЃЌB3-RAFБЛМЄЛюЃЌВЂжБНгМЄЛюIIIбЧРрSnRK2sЃЌДгЖјдіЧПаВЦШЯьгІЁЃ