����
��Ϊֻ�й���������ܽ��м�����һ��������⡣��ʵ�ϣ�ʹ�ò�ͬ��ʽ�����ʣ����������ʣ�Ҳ����ʵ��������ʽ�ļ��㡣�������ۼ������ѧ�ͺϳ�����ѧ֮���ЭͬЧӦ������ǿ���ϸ���������cellular computer�������Գ�Խͼ����㡣

����Ҫ��
1. ϸ��������ܹ����ض������ϳ�Խ�����������ⱻ��Ϊϸ����Խ�ԣ�cellular supremacy������Ϊֻ�й���Ļ������ܽ��м�����һ��������⡣��ʵ�ϣ�ʹ�ò�ͬ��ʽ�����ʣ����������ʣ�Ҳ������������ʵ�ַ�ʽ��
2. ��չϸ���������������Ȥ���뷨�������ڸ���������ʵ��Ӧ�á�����ҽѧ����̬ѧ��������ʹ�ý�����ڵ���Լ�ϸ���������
3. ��ͬ���͵���������Ա���Ч�ز�����ʵ�ּ��㹦�ܣ��粼������ֲ�ʽ�㷨�����ǵ�DNA���༭�Ը�֪���룬���������ඨ��Ĺ����ָ���ṩ�������Щ���Ѿ�ʵ�ֵļ��㡣
���ۼ������ѧ������ѧ��ʮ����һֱ�����������������ѧ��ͼģ������ϵͳ�Ĺ����Է�չ����ģ�ͣ������Զ������˹���������ݻ��㷨��������ѧ������Ϊһ����������������ϵͳ�Ĺ��ܡ�[4] ���磬��������70�����������ѧ�Ҿ�ʹ�ò����������������أ���ʱ�ſˡ�Īŵ��Jacques Monod��д������һ�主�������Ե�����������������������������ʽһ������[40]
������Ϊ��Ϣ�����Ǽ������ѧ�ͷ�������ѧ֮���Ŧ������Ϣ���䴦��λ������������ĺ��ġ��ڼ������ѧ�У���������״̬����ͼ���֮��ļ���ģ�Ͷ�������δ�һ�������һ������ָ�����������ͬ���أ�����ϵͳ����ͼ1A�е�ϸ��ϸ������֪��������̼�������Ӧ���������ڲ�����������Ӧ��ͨ��ʹ�úϳ�����ѧ[6]�����ڿ���������ϵͳ��ÿ������ľ������ʣ�����༭��ϸ����DNA�Ը�֪�µ����룩���Ӷ�ʵ��ʹ���������ʱ����Ϣ�����豸��[9]��һ�������ĵ�ͻ�Ʋ���Ϊ��ͳ��������ﵽ��Ӧ���ṩ�˿����ԣ�����ս�˼���Ĵ�ͳ�����Լ��ɼ�������ݡ���һ������ʤ�ĸ����п��ܽ��������ѧ�����µ�����Ϊδ���Ľ����ͷ�����ƽ��·��
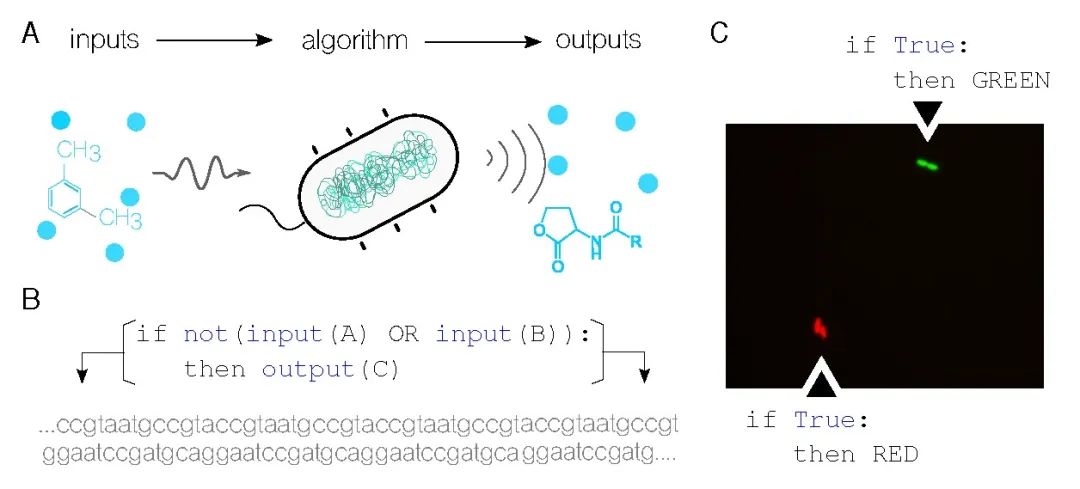
ͼ1. ���ϸ��ִ�����ﲼ����������A. ϸ�����и�֪����������ѧ���벢������DNA�б�����㷨��������������������������-�㷨-�����ϵ�Լ������������Ҫ��B. ��ϲ������������Լ��ɵ�ϸ���Ļ������С����磬����A��B�����ֻ�ѧ�����źţ�C�Ǹ����������ı������������¬��A �� B���� C ���Ա������DNA���У�Ȼ�������ϸ���С�C. ����ʾʾ���У���������ڼٵ�������ʵ�֣���ʹ��ӫ���������п��ӻ���������ϸ�������Ϊ����������ʲô��������ɫӫ�⵰�ף����ײ���ϸ�������Ϊ������ɫӫ�⵰����Ϊ���ǵ������
�������ϵͳ���̷�����ϵͳ���ڸ����ԵIJ��죬��Ϊ��ʹ�õ��������ʽ�Ȼ��ͬ������������ƺ����Ĵ�ͳ������ͬ������ϵͳͨ����Ȼѡ���������������ζ���������ڱ�̴�ͳ�����Ĺ��ߺͷ����ǿ�Ԥ������֪�ģ������书�ܵ�ʵ������ȷ�ı߽硣Ȼ����ϸ��������ͷ��ӵ��������ʲ����������˹������������й�����δ����ȫ��ʾ���������ǿ���ͨ����������ʵ�����粼�����ļ���ģ�ͣ��������ӵļ�����Ȼ������ս��Ϊ���ƶ����������ķ�չ��������Ҫ������Է���ϵͳ����ѧ�����⣬����չ������������ϵͳ�����ͼ������ۣ���Խ��ͳ��ͼ�������ģʽ��
��������ѧ�����½�չ������ƶ����������ķ�չ��������Ľ�չ�ɷ�Ϊ�ĸ��ؼ��Ρ��������20����90����������۽�������˼�������˸�����ķ�չ[17]����Ϊ��ʱʵ������ļ�����δ��λ�������һ�η�չʱ�ڣ�ֱ��20����90���ĩ����������ѧ��������㶼ȡ����������չ���ٳ���DNA���������ʵ�顣1994�꣬Leonard Adleman ������һ����б�־�������ʵ�飬ͨ��ʹ�÷��Ӳ�����DNA�������һ�����ܶ�·�����⣨Hamiltonian path problem, HPP����[3] �����η�����20����90���ĩ��21���ͳ������źϳ�����ѧ�ij��֣�������ϸ����ʵ�����š����غͷ�����·��[1,2,3]���Ľδ�2000���ĩ��ʼ����¡��DNA�ϳɼ���ȡ��������������ʹ�þ�ȷ�Ŵ������Ĵ�����Ϊ���ܡ�[39]ֵ��ע����ǣ��������۷�չ�����ʵ������������20����90���֮ǰ�������Ȼ�෴����һת��ǿ���˱��ĵĺ��Ĺ۵㣺���ۼ������ѧ�ͣ��ϳɣ�����ѧ�Ľ����ںϣ�Ϊ�����µĿ������ṩ�˶��صĻ��ᡣ
�����һƪ�����У����Ǵ����ˡ�ϸ����Խ�ԡ���һ����[29]����ָ��ʵ�����ض��������ܹ�ʤ������������ϸ���������������һ����������Խ�ԣ�quantum supremacy�����Ƶĸ����Щ����̫�������ڹ��������ó��������ر����ڸ�Ч�����ѧ���ⷽ�档Ȼ��������һЩ���������������������õ��������磬����һ���������⣬�������ֻ�����Ⱦ��x��y����϶�ֲ����̬ϵͳ��ɵ�Σ�ա�����������£������ڸ���̬ϵͳ�ڵ�ϸ������������ṩ������������Ҹ���Ҫ���ǣ�������ǿ϶��ģ���������Ϊֲ���ṩ���ȴ�ʩ�������ļ����豸������Ӧ�������������������Ƴɸ������ƹ��ܵĶ������ơ����Ƶ���������������Ҳ���������ҵ���������ҽ�����⡣
���ĶԹ���ϸ��������Ļ���������зֽ⣬������������˷ܵĸĽ�������
1. ʵ�ֻ�����������
�Ժϳ�����ѧ����������ʵ�ֲ����������ѳ�Ϊ����������Ҫ���㡣�����е����ģ���Ϊ���κθ���ʱ���ڣ������������ǻ�Ծ�ģ����������ǻ�Ծ�ģ��Ӷ�ʹ�����ܹ����������ֵ����/�ػ�1/0�����⣬��ʵ���������ѧ�Ƕ��������������������ף����Ҽ�������ʱ�ɻ�ȡ�ġ�[41]��ˣ�ʵ����ӱ���������ָ��Ļ�����ʲô�أ�
����ϵͳ����Ϣ�Ķ�����һ���������۵Ļ��⡣[2,44]Ϊ����������Ǽ���ǻ�Ծ��Ϣ�Ի������ʽ�洢��DNA�С����仰˵��ϸ��������������Ĺ��ܱ���������������У�����Щ�����ڱ�����֮ǰ���ַǻ�Ծ��ֵ��ע����ǣ����������Ϣ�ĸ������������ϵͳ���õ���Ϣ������DNA���ݴ洢��������Ϣ�Ķ��岻ͬ��[16]��DNA���ݴ洢�У�Ŀ���ǽ���������Ϣ�������κ�����ϵͳ��û���õ���Ϣ����������������У�����DNA��Ϊ����洢�豸�������������ָǰ�ߵĶ��塣
��һ��������ʱ������������ʣ�ִ����Ϣ�б���Ĺ��ܡ����仰˵�����DZ�����������ִ���书�ܡ����������ѭ����������������Ը�����Ҫ�رա����磬���빦��C�Ļ���ֻ�����ض�����A��B���ʱ�Żᱻ�����ͼ1B��ʾ������������У����б��������Ա�����ģ����롢����Լ��������ǵ��㷨�����Ա����塣
ͼ1Cչʾ������Ϊֹ������ϸ���ٵ�������ʵ�ֵ����������[22]����ʹ��ӫ�����������ͼ��չʾ�������е�ϸ��������������Ƭʱ��ʵ�鿪ʼʱ��������ϸ�����շ��ѳ��ĸ���������ϸ��������������ɫ�źţ����������������ڱ������ɫӫ�⵰�ף��ײ���ϸ��������ɫӫ�⵰�ס�������ϸ����ѭ��ͬ������������������Σ������Ӧ�ñ��������������£�û�ж������룬��ʹ���Ϊһ������䣩��ֵ��ע����ǣ�����������DNA�����У�û����Ұ������������Ȼ���֡��෴�����Ѿ�����Ϊ���ӵ����ǵĻ������С�
��20����90���ĩ�������о���Աһֱ��ϸ��[54]����ĸ[15]�Ͳ��鶯��ϸ��[7]��ʵ���Ŵ��� NOT �������������������š�����������Щʵ�ַ�ʽ�Ѿ��Ը��ַ�ʽ�õ��Ľ������磬�ű���Ƶø�ģ�黯���Ա����ǿ���������һ��������Ŵ���·������Ҳ��ø��ӱ������õر�����ʹ��ƹ��̸����Զ��������⣬��ѧ��̽���˸���ϸ�����ƣ�����չ���ڹ�����Щ�ŵĹ��߰���������תø��ת¼��Ӧ��������ϸ��Ⱥ��[26,46]�����ǵ���ϸ������ϸ��������ļ�������������ϵͳ��DZ�����бȽ��Ǻ���Ȥ�ġ���ϸ�����ܹ�ʵ�ֵ�һ���൱������������������Ҫ6��8�������˾�ϸ����4000����������������ֻ�Ǹոտ�ʼ��������ϵͳ�ļ���������������ˣ�������ϸ������ƺ�ʵ�ֲ�������������Ѿ���Ϊ������̡�
Ȼ����������״̬����ͼ�����ȣ��������һ�����ļ���ģ�͡�������ϵͳ����Ϣ�������滹�и�����ṩ�ġ���ô��������Щģ���Ѿ�ʵ���ˣ�������Щģ���д�̽����
2. ���������ģ��
����ģ����һ�����۸�����������˽����ض������¿��Խ��ʲô�����Լ���ν�������Ƕ����ִ��ɵ��������Ĺ��������ķ�չ������Ҫ��Ȼ��������������У�������Ҫ�Բ�ͬ��ʽ�����������������������Լ����㷨�����DZ���Ū������������������������Բ�ͬ�ķ�ʽ������Ϣ����һ��ս�漰�ӻ�����������ӵ�·�ıȽϣ����쵽���Թ������������ıȽϡ�[31]
�������ǶԴ˵������в�ȫ�棬������ӵ�й㷺����������ԭ�����������������ָ������Ԫ�ص�����������������������豸������������ϸ����ʵ�ֱȼIJ����������ӵļ���ģ���Ǻ����ģ����о���Ա����һ�����Ѿ�ȡ���˽�չ��һ����ϣ�����о������ǹ���״̬������Щ�豸�ܹ����ض��������Ӧ�£�������������״̬֮�����ת����
��������ֻ���֮һ�ǻ����л�����[23]����2000���ڴ˾��й�������������������ȶ�״̬�����ΪA��B����ͼ2A��ʾ���Լ��������룬I1��I2��������������״̬���л������豸��ÿ��״̬ʹ�ò�ͬ�ı���ϵͳʵ�֣���һ������������������У��������ǿ��Ը�����Ҫ�ⲿ���ӵ�С���ӡ�����ʱ�𣬻����л������Ѿ�������汾���Ż����Լ����������������ܵ������о���[42]
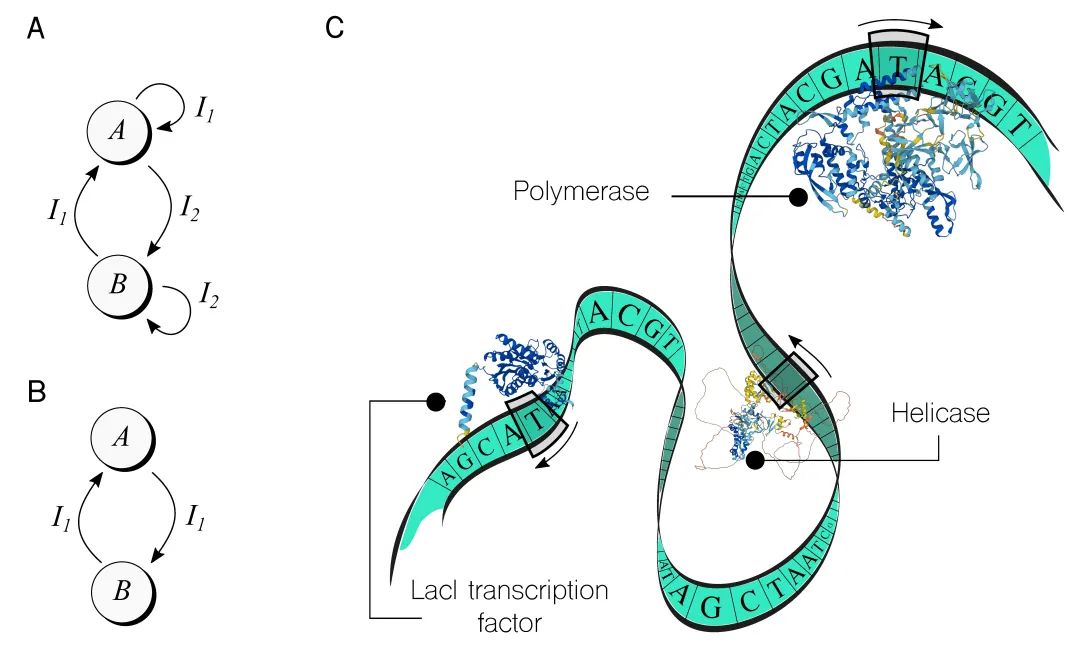
ͼ2. ��ϸ��������ʵ�ּ���ģ�ͣ���Խ�������A. ��Խ�IJ�������̽��������״̬������ǿ��ļ���ģ�͡����磬�����л�����ʹ�����ֻ�ѧ���루Ix�����ȶ�״̬A��B֮���л�����ʾ��ͼ��ʾ��B. ʹ�ý�һ���������ı�״̬�Ļ��ظ�����ս�ԣ����Ѿ�����ʾ����C. ����DNA������һ������ֽ�������ݴ洢�����Ա�ͼ��ʽ����ϵͳ�е�ø��ȡ�ʹ������������ϵͳ��δ��ȫ�����⣬Ҳû�б���ʽ��Ϊ�����豸������Ϊ��ʾ����ϵͳ��������Ϣ���������ṩ�˼������ĵĿ����ԡ�����һ�����£�һ��ֵ��ע��������ǽ�����һ��ø��װ�ø��Ϊ�ܹ�ִ�������溯���ļ���ϵͳ[10]����һ��������Ѿ�������ʵ�������[50]����������AlphaFold���ɵ����ʽṹ https://alphafold.ebi.ac.uk����
��һ��ֵ��ע����������ƿ�-�ƹؿ��أ���ͼ2B��ʾ��Ҳ�ڴ˾�ϸ����ʵ�֡�[34]����״̬��ֻ��Ҫһ�������źž����ڲ�ͬ״̬֮��ת��������ͨ�����Ӷ���Ŀ����ź�������ʵ�ֵĸ����ԡ���ˣ�ϸ����һ���ⲿ�̼������˿�����Ӧ��
����ð�վ����������㳢�Գ�Խ��ͳģ�ͣ������Ŵ�����ʵ�����������ļܹ���[33]�������ܹ�ʹ��ģ�����Ϳ����Խ��������˹��������֪��[43]������汾����Щ������Ȼȡ���˳ɹ������ҿ��ܸ��ӽ���Ȼ�㷨�������ܻ��������������������ģ���д����֡���ʵ�ϣ�һЩ�о���Ա����뷨������������̳�Խͼ����ľ����ԣ�[36]��������ϵͳ��δʵ��ʵ�֡�
����DNA��һ��ͼ���������������У�DNA����ͼ����е�����ֽ��������д�ŷ��š�Ȼ������DNA������£�ֽ�����������ַ��ţ�A��C��G��T��������ͷ����ø������ֽ��˫������ȡ��д���ִ����������ģ���ͷͼ�������Щͷ���������첽���������ǵ���Ϊȡ�����ض������ָ���Щ�����ָ���ÿ��ͷ�����Ƕ��صġ����⣬���Dz�������DNA������������з��룬��ɢ��ϸ�����У�Ȼ�����½�ϵ�ֽ���ϲ�����������
��ͼ2C�У�һ����ͨ��DNA��������һ��ͼ�����ͼ��չʾ����������ϵͳ���ڶ����ͷ�������ض����͡����磬�ۺ�ø��һ��ø������ȡDNA��ֱ��ʶ���ض��ַ�����Ȼ������һ����������ķ��Ŷ�Ӧ��RNA���ӣ�ֱ������ֹͣ�ַ�����������̳�Ϊת¼������ø����һ�����͵ġ�ͷ������ʶ��һ����Ϊ���������ַ�����Ȼ�������ļ����ϸ���������� DNA���Ӷ�����ͼ�������е�����ֽ����ͼ2C����ʾ�ĵ�����ͷ��LacIת¼���ӣ�TF����ת¼�����Ǻϳ�����ѧ���߰��Ĺؼ����֣�[27]���ڴ�������ֹ�ϳɻ�����ж�����ͨ��һϵ�й����Ի����豸������/��ģʽ��
��������ͼ�����һ����Ȥ�����ǣ����Ĵ�Ԫ�ض����Ա���̡����磬ֽ���ϵķ��ſ��Ա��༭��ת¼���ӵĴ��ڻ�ȱʧ���Ա����ڣ����������ֻ�����������������ģ��[19]������̬[47]���㷽���ij��֡��������֡�ͷ������ۺ�ø���Ĺ���Ҳ���Ա��༭�����磬�ۺ�ø���Ա�����Ϊ�������ض�������ϣ����Ѿ��������ͷ�������ڼ���֮�С�
��Ȼ��DNA��Ϊͼ����ĸ���ȷʵ������ʤ��������Ȼ�Ƕ�����ϵͳ�����ԵĹ��ȼ�Ȼ�����������Ϊ����������ļ��������Լ�����������ִ���ض������ṩ��һ�����õĿ�ܡ����磬ͨ���о��������������ƣ����ǿ��Է����µļ���ģ�ͣ���Щģ�Ϳ��������������ܱȵ�ǰ�����������ָ��õ�ϸ���������������ϸ����Խ�Եķ���[29]
��ĿǰΪֹ�����IJ����ڼ������ѧ���������Ƕȿ��������Ϣ�ķ�����Ȼ����ϸ�����㲻���漰���������漰�������������ʵ�֣��漰Ӳ�����⣬���Ҳ�漰��������[5]
3. Ӳ�������������߶�����
����Ӳ�������������ṩ���õĸ���������ʽ������ϵͳ���ܵı������ڽ����С�[18]Ȼ����ֵ��ע����ǣ��ϳ�����ѧ�ڹ��������豸ʱ�����ڹ��̺ͽ�����ȡ���ˣ�Ӳ�����ͨ��ռ��������λ��������һ�������������£�ƫ���˼����������㹤�̡�
��Ȼ��ѧ�������������ŵ�ʵ������������֮���������߸�ֲ�ڵ���������������Ϊ��������ṩ�����õĸ���ͳ���ͬʱҲ�������ԡ����磬��������Ϊ�������Ŀ�ܣ������˲����ݱ�������ṹ�����⡣��Ӳ����ͬ���������ʡ������Ϊʪ����������ݲ��ϱ仯�Ļ���������Ӧ��ͻ�䡣�������һ���ش���ս�����������Դ˽������ۡ���Ȼ���������ģ�飨������ϵͳ�ͷ������ӣ�������������Թ�������ĵ�·��������ȱ�������������ԣ����ǵ���ϵͳ��һ���ؼ������������Ѿ���Ŭ������ʪ��[8,38]�����̿��ܽ�����һ�����dz������õĸ����ˣ����������ʵ����Ϣ�����ϣ������뵽����������ṩһ�����ʺϲ�������ϵͳ�Ŀ�ܡ����磬�������Ų���ѭ�����豸�г���������/���������TTL��CMOS��ÿ�����Ŷ����ܳ��ֲ�ͬ������/���ģʽ������ÿ���ŵĶ�̬��Χ��ʶ������ŵ�Ψһ���������⣬��ѭ��������ԭ�����·ͨ���ǰ�˳�����еģ��������ǵĵ��Ӷ�Ӧ��һ����Ȼ������Ȼ�Ŵ������и�������ͷ�����·������ܾ����㷨���ƣ��������뵽������õķ���������Խֱ����ǰ����ơ�[30]
��Ϊֻ�л��ڹ���������ܽ��м�����һ��������⡣��ʵ�ϣ���Щֻ����ѧ��ʽ�����ض�ʵ�֡�ʵ���ϣ�ʹ�ò�ͬ��ʽ�����ʣ����������ʣ�Ҳ����ʵ��������ʽ�ļ��㡣
��ϸ������������У����ǿ����ڸ���������Ӳ��������������ϸ�����㷽������һ�ַ����漰�ֲ�ʽϸ�����㣬[24,26,46]����ϸ��Ⱥ����ʵ�ָ������ܡ�Ⱥ���е�ÿ��ϸ��ִ���ض��������ܣ�Ȼ������Ϊ���봫�ݸ�����ϸ�����������Խ����ֽܷ�Ϊ������֣�����������ϸ����ϸ�������䵱Ӳ���ṹ�����Ŵ�������䵱������ͨ�������ֽܷ�Ϊ������ֲ������ϸ�������Ը��õع�������չ���㸺������Ϊϸ������������Դ���������ڶ��ַ����ǿ���������[25]���漰����ʵʱ�������õĻ����豸�����磬һ����������ܹ����ݿ����źţ����κ�ʱ��ִ�ж��Ԥ�����������е�һ�֡�[13]���ַ������ӽ����Ƕ����������⡪����Ϊ���������õĹ��Ӳ����Ȼ��һ����ս��������ʹ�豸����������Ӧ��ǿ���书�ܿ��Ը�����Ҫ���и��ġ�
�����漰��ȷ���ݻ����ϸ������Դ������������·�����¶��Ϸ���֮�⣬������̽�������϶��µķ�����[45]��Щ����ּ��ͨ���ı�����̼�������Ҫ��ϵͳ����������͵���ԭ��ϵͳ�������ѱ�Ӧ���ڲ����ؼ���������㹦�ܣ�����䣬[11]���������±�̿��Ʋ��Զ����������������Ӳ������һ�뷨�������뵽���������á�
���������������������Ӳ���������ı����С���Ϊ��ȷʵ��һ����Ҫ�ĸ���������Ȼ���߶����ж��ص����ƣ�����ʵ�ǣ�����ϵͳ�Ƕ�һ���ġ����ǼȲ���Ӳ��Ҳ���������������ٲ������Ƕ�������������������ۼ�������չ�ֵ�Խ��֤��
4. �����ԵĹ���
���漰��ϸ������ʱ���������ѧ����ϵͳ����������һ��Ȼ���ҵ���ͬ������һ���ش���ս��һ���棬�������ѧ�漰�㷨�����Ժͽ����������ѡ�Ȼ����Ӳ�������Բ�����һ����Ҫ��ע�㣬��Ϊ�����������ϵͳ�ǿ�Ԥ��ģ�û�����벻�������ԡ���һ���棬����������Ӧ���ڷ�������ϵͳ�����ϸ�����漰�������֮�������ò�����ӿ�����Ժͷ���������[51]����Щϵͳ���ڶ��䶯̬������ȫ����IJ��������Ԥ�⡣
��������漰�����������������ǵĽ�����Ȼ����ȷ��һ��ϸ����һ������ϵͳ��Ϊ������ƻ����豸��Ҫ���ǿ��Խ��������ͽ�������ĸ����ԡ�
�Ӹ���ϵͳ�ĽǶ�������ͼ3A������Ҫע����ǣ�һ���Ŵ������DNA���в�����Ӱ���ض�����ʵ�ֵ�Ψһ���ء����ֲ�������˸����ԣ��������ϳ����з�����ϸ����Ⱦɫ��λ�ã�����ܻ��ԭʼ������������Ա仯�����⣬Я�������ϸ�����̣��Լ����ֺ;��꣬�����Ըı�ԭʼ���������������˵��������������[53]����һ����������������������������⣬ϸ�����ǹ��������ģ��������и��ӽṹ������õ���ȺҲ����ԭʼ���������̬������Ϊ������������붯̬�仯����Щ�仯������ǰԤ�⡣[49]�����������豸ּ�ڽ���뻷����ص����⣬��һ������Ϊ��Ҫ������ϵͳ�����Զ������������˾���ս����Ӧǿ�������ս���Dz�����Խ������ϵͳ��������������ݻ����Ծ��˵ľ��ȸ�Ч��ִ������ͬʱ���������ԡ���Ȼ���Ⲣ����ζ��������Դﵽ��ͬˮƽ�������������ʣ������ǿ϶����Ա�Ŀǰ���ø��á�
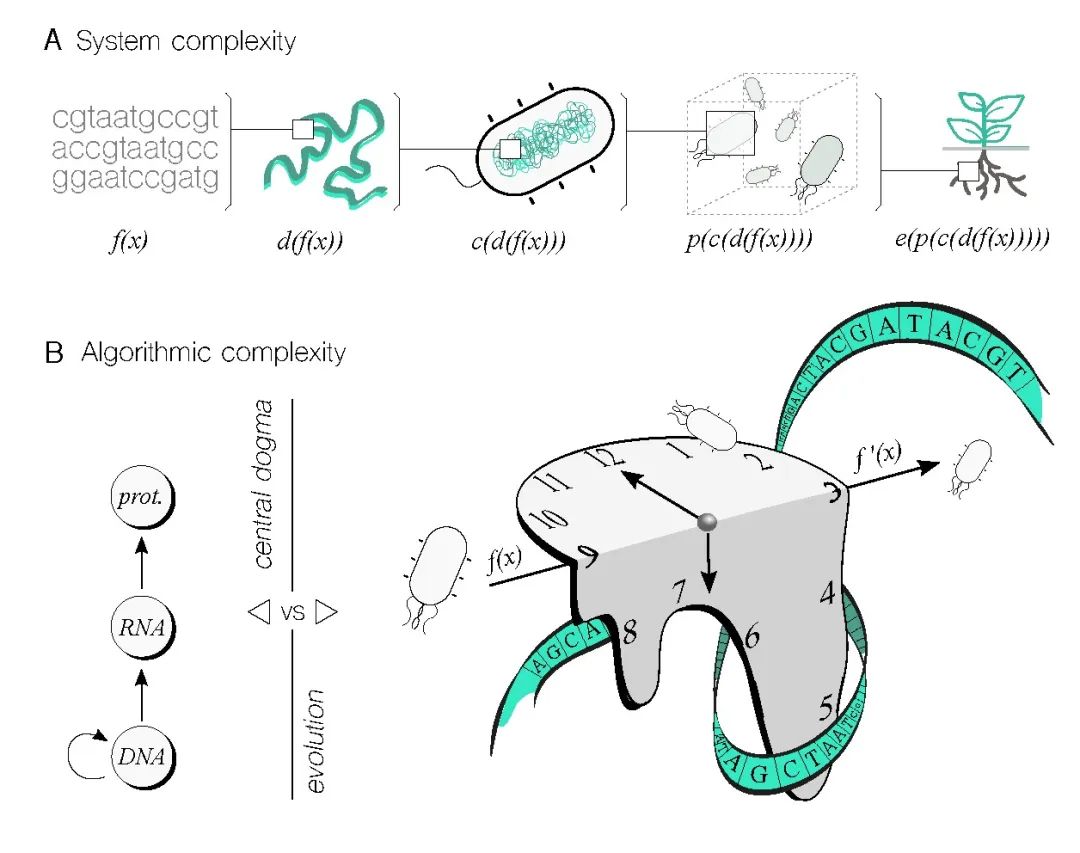
ͼ3. �ڸ���ϵͳ�ͼ������ѧ��Ѱ��ϵͳ���㷨������֮��Ĺ�ͬ������A. ��ϸ���Ǿ���ӿ�����Եĸ���ϵͳ����Щ��������ɲ���֮�������õ��¡�Ϊ����DNA������ʵ�ֹ��ܣ���Ҫ���Dz�ͬ��ε���Щ����ã���Ԥ����������豸�����ܡ���ͼ��ʾ�����뵽 DNA�еĺ��� f��x������ͨ��������λ�á����������塢��Ⱥ����ѧ����̬λ�����ؽ����ģ�������Щ���ض������˺������ܵĸ����ԡ�B. �㷨�����Զ����κμ��㹤����������Ҫ��Ҳ�����ڲ�ͬ���������̡����ܷ�������ѧ�����ķ���DNA��RNA�ٵ������ʣ���һ�����ֱ�ӵĹ��̣��������������ô������ڻ�ʱ������ʾ�����и����ӵ���Ϣ����������һ����������ͨ���Կ��ŷ�ʽ��Ӧ���ϱ仯�Ļ������ݱ���µĺ���������������Լ��������ã��㷨������������ش���ս��
��������豸���㷨������ֵ�ø����ע����Ϊ���Ǽ������ѧ�е�һ�����Ļ��⡣���漰�ش�һ�����⣺���ض�����ϵͳ�У��������룬�������ж����ѣ����ڽ���������ij�������������ʱ��Ϳռ���������������£����һ������ϵͳ�ﵽ��������ʱ�����һ��������ôǰ��������ʱ���ϱ���Ϊ�����ӡ����������������ֲ��䣬��Ϊ�������������ײ��㷨ϵͳ�ĸ����ԡ������ǽ�ʱ����Ϊ����������Ч���������Բ���������Ҫ����Ĺ۵㡣������̵ĸ����Կ���ֱ�۵�����Ϊ�ڽ��������ṩ�������������в�ͬ����ͼ3B�������磬��������ѧ�����ķ���CD�������˻��������̣�������DNA�������ʣ��漰ת¼�ͷ��룬��ֻ�ܽ����DNA�ͱ��������ϸ���������ڵ�������⡣ͨ�������ķ�����Ϊ�㷨�����ǿ��Լ������������ʱ�䡣�����뻯��ת¼�����龰�У�����һ���ڵ�ĵ��ײ����յ���������һ������ı��ʱ�佫�漶�������������ӡ�Ȼ��������ʱ�临���Կ϶����ܵ��������ơ����ӷ����ԡ���Դ������������ص�Ӱ��������仯��
��Ȼ���������һ����Կ��ٺͼĹ��̣�������Ҫ���ӵöࡢ���۵öࡣ�������Խ�����㷺�����⣬���㷨��������Ȳ�ֱ��Ҳ�����١�����ϵͳͨ��������Ӧ��������������Ϊһ���㷨������ӡ����̵ġ�ͨ��ͻ���ѡ���辫ȷ�����ݻ����̵�ʱ�临������һ���ش���ս��Ȼ����ʵ����һĿ�����ɽ���ʾ���صĻ���ϸ�ڣ��Ӷ������������������㹤������Ȼ�ݻ������ܵ���Ȼ�ݻ���������������δ��ȫ���临���ԺͿ����ԡ���ˣ�������Ȼ�ݻ��ĸ�����������������㽫������ġ�
���൱ǰ��Ŭ��ּ�ڿ˷��ݻ���DNA����IJ��ɱ����Ӱ�죬����ܵ���ͻ�����ɥʧ��[48]Ȼ��������Ӱ�족���������ܻ�Ӿ����⣬��Ϊ�ݻ����������濴��������Ϊ�ƻ����ܳ���Ĺ��̡��෴�����ݻ���Ϊһ�ֹ��߿�����DNA������������Ϣ����������ʹ���ܳ�Խ��ǰ���ơ��������ݻ����Գ�Ϊһ����������ѣ�������һ���ϰ���
���ܴﵽȫ������ϵͳ�����ԣ�ͼ3A������������ÿһ��ϸ���ƺ���̫���ܣ������ǿ�����Ч�ؽ�ijЩϸ�ڳ���Ϊ����ģ�顣���ַ���ʹ�����ܹ�����ģ��������������ѧ�������������������ڲ����������磬�Ŵ�����ģ�飨ͼ3A�е�f��x������װ���������ϸ�ڣ����DZ�����������ϸ�ڲ����������ǹ���������ȫ�Ļ������š�
�������Ӱ����ڲ���ȫ������������������[53]�������ʹ�û������ţ�ͼ3A�е�c��d��f��x������������ʹ����Ⱥ����ѧ����ȫ�˽�Ҳ��ʵ����Ч�Ķ�ϸ���ֲ�����[35]��ͼ3A�е�p��c��d��f��x�������������⣬�ݻ�����ȷʵ������������Ŵ���·�IJ���[56]����ʹ����Щ���̵���������������������
�ܵ���˵������ Donella Meadows �Ļ���������������[����]ϵͳ��Ū������ǣ������ǿ��������ǹ��裡��ÿһ�ζ���Щϵͳ���¶��춼��ʵ�ָ��Ƚ���������㣬���ø����ԵĻ�����Ȼ���㣬��ʹ���Dz�δ��ȫ��������
5. ��ս�����
�Ľ�ϸ��������ı����һ�������ս�Ե�������Ҫ��ǿ���ۼ������ѧ�ͺϳ�����ѧ�Ľ��档���磬ͨ����ʽ���µļ���ģ�ͣ����ǽ���������Ȼ��̬ϵͳ�����⣬����ǿ�ϳ��Ŵ����ܵķ�Χ��Ȼ����������������ѧ�����µļ���ģ�������ѵģ���Ϊ����ϰ���ڵ�ǰ���ڹ��ʵ�ֺ����е���ѧ�����⣬����ϵͳ���㷨������֮����ص����Ե����������������ʵ����ͼ���ģ�͡�
��������һ���ؼ���ս����Ϊ��ǰ�ϳɹ��ܵ�ʵ�������ܿ����ݻ�����ѧ����������Ϊ���Dz�֪����ζ�����й������⡣[14,29]���ڴ���Ӳ����е����Ĺ���ѧ��ʱ��Ϊ������ս�ԡ�Ȼ���������������̽��жԱ�ʱ�����ֱȽϾͲ���ô������ս���ˡ�[12] ��ʵ�ϣ��������ѧ����֧�����ֹ۵㣬���ǵ���ʮ��ǰ������ѧ�л����е��ݻ����㣬��һ����Ȥ�ķ����dz�Խ������صĹ����䡣ʵ���ϣ���ϸ��Զ��ֹ�ڴˡ������³´�л������ϸ���Ĺ��̿��Ե��¸������ڻ�ϸ��ʵ�ֹؼ��������̵����μ���[28]��
��չϸ�������������ѧ���Ͼ���Ȥζ�ԣ������ڸ����������ʵ��Ӧ�ã���ҽѧ[55]����̬ѧ[21]��������ʹ�ý����Ѿ����ڵ���Լ�ϸ�����������ЩӦ�����������һ���ص����������Ҫ��������Ľ�����������⣬������̬������Ϣ�����Ⱦ���̽��������ͼ��[20]���о���������Ҫ�������һ��ͻ��������ͻ���˷�������ѧ����Ϣ��������֮��Ľ�����ϵ��[37]��Щ�ض���ս�ƺ����ʺ�������㷽���������Ǵ�ͳ�ļ��㷽����
δ�����и�����ս���������ඨ���ϸ������������Ľ����в��������ǰ���ǹ����ġ����磬���������������ڵģ����ҶԴ����ǵ����嵽������̬ϵͳ�ȸ���ϵͳ��������Ҫ�������棩���á�[21]����Ļ�����ǰ���ȴ�������Χ�������ϵͳʧ������ʱ�����±����Щ����������ƽ��ϵͳ���������ʣ����������һ��ֵ�������ϣ�����о�����
�����
1. Abelson, H.et al. Amorphous computing. Commun. ACM 43, 5��May 2000��, 74�C82.
2. Adami, C. The use of information theory in evolutionary biology. Annals of the New York Academy of Sciences 1256, 1��2012��, 49�C65.
3. Adleman, L.M. Molecular computation of solutions to combinatorial problems. Science 266, 5187��1994��, 1021�C1024.
4. Amos, M. and Goñi-Moreno, A. Cellular computing and synthetic biology. Computational Matter��2018��, 93�C110.
5. Andrianantoandro, E., Basu, S., Karig, D.K., and Weiss, R. Synthetic biology��New engineering rules for an emerging discipline. Molecular Systems Biology 2, 1��2006��, 2006.0028.
6. Ausländer, S., Ausländer, D., Fussenegger, M. Synthetic biology��the synthesis of biology. Angewandte Chemie Intern. Edition 56, 23��2017��, 6396�C419.
7. Ausländer, S. et al. Programmable single-cell mammalian biocomputers. Nature 487, 7405��2012��, 123�C127.
8. Beal, J. et al. The long journey towards standards for engineering biosystems��Are the Molecular Biology and the Biotech communities ready to standardise?EMBO Reports 21, 5��2020��, e50521.
9. Benenson, Y. Biomolecular computing systems��principles, progress and potential. Nature Reviews Genetics 13, 7��2012��, 455�C468.
10. Bennett, C.H. The thermodynamics of computation��a review. Intern. J. Theoretical Physics 21,��1982��, 905�C940.
11. Biswas, S., Clawson, W., and Levin, M. Learning in transcriptional network models��Computational discovery of pathway-level memory and effective interventions. Intern. J. of Molecular Sciences 24, 1��2022��, 285.
12. Calcott, B. et al. Engineering and biology��Counsel for a continued relationship. Biological Theory 10,��2015��, 50�C59.
13. Canadell, D. et al. Implementing re-configurable biological computation with distributed multicellular consortia. Nucleic Acids Research 50, 21��2022��, 12578�C12595.
14. Castle, S.D., Grierson, C.S., Gorochowski, T.E. Towards an engineering theory of evolution. Nature Commun. 12, 1��2021��, 3326.
15. Chen, Y. et al. Genetic circuit design automation for yeast. Nature Microbiology 5, 11��2020��, 1349�C1360.
16. Church, G.M., Gao, Y., and Kosuri, S. Next-generation digital information storage in DNA. Science 337, 6102��2012��, 1628�C1628.
17. Conrad, M. Molecular computing. Advances in Computers, 31 . Elsevier, 1990, 35�C324.
18. Danchin, A. Bacteria as computers making computers. FEMS Microbiology Reviews 33, 1��2008��, 3�C26.
19. Daniel, R., Rubens, J.R., Sarpeshkar, R., and Lu, T.K. Synthetic analog computation in living cells. Nature 497, 7451��2013��, 619�C623.
20. de Lorenzo, V., Marliere, P., and Sole, R. Bioremediation at a global scale��from the test tube to planet Earth. Microbial Biotechnology 9, 5��2016��, 618�C625.
21. De Lorenzo, V. et al. The power of synthetic biology for bioproduction, remediation and pollution control��the UN��s Sustainable Development Goals will inevitably require the application of molecular biology and biotechnology on a global scale. EMBO Reports 19, 4��2018��, e45658.
22. Espeso, D.R., Martçnez-Garcça, E., De Lorenzo, V., and Goñi-Moreno, Á. Physical forces shape group identity of swimming Pseudomonas putida cells. Frontiers in Microbiology 7, 1437��2016��.
23. Gardner, T.S., Cantor, C.R., and Collins, J.J. Construction of a genetic toggle switch in Escherichia coli. Nature 403, 6767��2000��, 339�C342.
24. Goni-Moreno, A., Redondo-Nieto, M., Arroyo, F., and Castellanos, J. Biocircuit design through engineering bacterial logic gates. Natural Computing 10,��2011��, 119�C127.
25. Goñi-Moreno, A. and Amos, M. A reconfigurable NAND/NOR genetic logic gate. BMC Systems Biology 6, 1��2012��, 1�C11.
26. Goñi-Moreno, A., Amos, M., and de la Cruz, F. Multicellular computing using conjugation for wiring. PLoS One 8, 6��2013��, e65986.
27. Goñi-Moreno, Á., Benedetti, I., Kim, J., and de Lorenzo, V. Deconvolution of gene expression noise into spatial dynamics of transcription factor�Cpromoter interplay. ACS Synthetic Biology 6, 7��2017��, 1359�C1369.
28. Goñi-Moreno, A. and Nikel, P.I. High-performance biocomputing in synthetic biology�Cintegrated transcriptional and metabolic circuits. Frontiers in Bioengineering and Biotechnology 40,��2019��.
29. Grozinger, L. et al. Pathways to cellular supremacy in biocomputing. Nature Commun. 10, 1��2019��, 5250.
30. Grozinger, L. and Goñi-Moreno, Á. Computational evolution of gene circuit topologies to meet design requirements. In Proceedings of the 2023 Artificial Life Conf. MIT Press, Cambridge, MA, USA.
31. Jonas, E. and Kording, K.P. Could a neuroscientist understand a microprocessor? PLoS Computational Biology 13, 1��2017��, e1005268.
32. Knight, T.F. and Sussman, G.J. Cellular gate technology. Massachusetts Inst of Tech Cambridge Artificial Intelligence Lab, 1998.
33. Li, X. et al. Synthetic neural-like computing in microbial consortia for pattern recognition. Nature Commun. 12, 1��2021��, 3139.
34. Lou, C. et al. Synthesizing a novel genetic sequential logic circuit��A push-on push-off switch. Molecular Systems Biology 6, 1��2010��, 350.
35. Mac��a, J., Posas, F., and Sol��, R.V. Distributed computation��The new wave of synthetic biology devices. Trends in Biotechnology 30, 6��2012��, 342�C349.
36. MacLennan, B.J. Natural computation and non-Turing models of computation. Theoretical Computer Science 317, 1�C3��2004��, 115�C145.
37. Manicka, S. and Levin, M. Minimal developmental computation��a causal network approach to understand morphogenetic pattern formation. Entropy 24, 1��2022��, 107.
38. Mart��nez-Garc��a, E. et al. SEVA 4.0��An update of the Standard European Vector Architecture database for advanced analysis and programming of bacterial phenotypes. Nucleic Acids Research 51, D1��2023��, D1558�CD1567.
39. Meng, F. and Ellis, T. The second decade of synthetic biology��2010�C2020. Nature Commun. 11, 1��2020��, 5174.
40. Monod, J. Chance and Necessity��An essay on the natural philosophy of modern biology , 1971.
41. Nielsen, A.A. et al. Genetic circuit design automation. Science 352, 6281��2016��, aac7341.
42. P��jaro, M., Alonso, A.A., Otero-Muras, I., and V��zquez, C. Stochastic modeling and numerical simulation of gene regulatory networks with protein bursting. J. Theoretical Biology 421,��2017��, 51�C70.
43. Pandi, A.et al. Metabolic perceptrons for neural computing in biological systems. Nature Commun. 10, 1��2019��, 3880.
44. Pattee, H.H. and Rączaszek-Leonardi, J. How Does a Molecule Become a Message? LAWS, LANGUAGE and LIFE��Howard Pattee��s Classic Papers on the Physics of Symbols with Contemporary Commentary. Springer, 2012, 55�C67.
45. Pezzulo, G. and Levin, M. Top-down models in biology��Explanation and control of complex living systems above the molecular level. J. The Royal Society Interface 13, 124��2016��, 20160555.
46. Regot, S. et al. Distributed biological computation with multicellular engineered networks. Nature 469, 7329��2011��, 207�C211.
47. Rizik, L. et al. Synthetic neuromorphic computing in living cells. Nature Commun. 13, 1��2022��, 5602.
48. Sleight, S.C., Bartley, B.A., Lieviant, J.A., and Sauro, H.M. Designing and engineering evolutionary robust genetic circuits. J. Biological Engineering 4, 1��2010��, 1�C20.
49. Sol��, R.V., Montañez, R., and Duran-Nebreda, S. Synthetic circuit designs for earth terraformation. Biology Direct 10, 1��2015��, 1�C10.
50. Srivastava, R. and Bagh, S. A logically reversible double Feynman gate with molecular engineered bacteria arranged in an artificial neural network-type architecture. ACS Synthetic Biology 12, 1��2022��, 51�C60.
51. Stoof, R. and Goñi-Moreno, Á. Modelling co-translational dimerization for programmable nonlinearity in synthetic biology. J. Royal Society Interface . 17, 172��2020��, 20200561.
52. Tas, H., Grozinger, L., Goñi-Moreno, A., and de Lorenzo, V. Automated design and implementation of a NOR gate in Pseudomonas putida. Synthetic Biology 6, 1��2021��, ysab024.
53. Tas, H. et al. Contextual dependencies expand the re-usability of genetic inverters. Nature Commun. 12, 1��2021��, 355.
54. Wang, B., Kitney, R.I., Joly, N., and Buck, M. Engineering modular and orthogonal genetic logic gates for robust digital-like synthetic biology. Nature Commun. 2, 1��2011��, 508.
55. Xie, Z. et al. Multi-input RNAi-based logic circuit for identification of specific cancer cells. Science 333, 6047��2011��, 1307�C1311.
56. Yokobayashi, Y., Weiss, R., Arnold, F.H. Directed evolution of a genetic circuit. In Proceedings of the National Academy of Sciences 99, 26��2002��, 16587�C16591.