�˹�������Artificial Life, Alife����һ�����˵Ŀ�ѧ���о�����������ͨ������ģ�ͺ�����ģ�⣬̽�������ĸ���DZ����ʽ��Alife������ͼģ����Ƶ�������֪��������ʽ�������Դ���ȫ�µġ����ܴ��ڵ�������ʽ���Ӷ���չ���Ƕԡ���������һ��������⡣ͬʱ��Alife��������������Ҳ��ΪAI�ķ�չ�ṩ��Ҫ��кͷ�ʽ��
������һ�����Ӷ����ص���������˵����What I can't create, I do not understand�������Ҫ������������õķ�ʽ���¾���ȥ���������������ǹ���̫��ħż��Golem������ϣ��������˹���й��ܳ�����ʦ��������ѧ��Ʒ�еĸ�����˹̹���ִ��ƻ�С˵�����ִ��������ij嶯��Խ��ͬ�������᳹����������ʷ�������죬���ִ��������ij嶯������Ϊ�����㹦�ܻ��������������ڿƼ�ʵ����չ����DZ�������������˹��������˹���������Ϊ���������������������д��ڵ�������������Ҫ���ס�
�˹�������Artificial Life, Alife����һ�����˵Ŀ�ѧ���о�������������ͨ������ģ�ͺ�����ģ�⣬̽�������ĸ���DZ����ʽ��Alife������ͼģ����Ƶ�������֪��������ʽ�������Դ���ȫ�µġ����ܴ��ڵ�������ʽ���Ӷ���չ���Ƕԡ���������һ��������⡣�����ʹ�ü����ģ���������̣��о�����֯ϵͳ���Լ�������ģ����Ȼѡ��ͽ������̵�������Ӳ��ϵͳ�����Ľ��ڻع�Alife��ʷ�Ļ����ϣ����������µ��о������ʵ����չ��
1. ���������˹�����
��ͳ����ѧ�У�����ͨ������Ϊ����DNA�͵����ʹ��ɣ������³´�л����ֳ�������Ŵ��ݻ���һϵ����״����Ȼϵͳ��ֱ��1944�꣬����ѧ��Ѧ���̷�����������ʲô����What Is Life?���������Ű����ض���������֯�ṹ������������Ϊһ������-��Ϣ��ϵĸ���ϵͳ������˼��Ȼ�Ӧ�˹�ȥ��Ҳӭ���˵��¡��������Ӵ�����л������ص����л��ۺͻ�е��֮�䣬ҲΪAlife�������о��ṩ�˿��ܡ�
����ͬһʱ�ڣ����ͬ����Ŀ�ѧ�ҿ�ʼ�˶�Alife��̽������һ�����̽�����¿��Է�Ϊ�����˳���
��һ�Σ�20����50-60��������Ը���̽����
�����ʱ�ڣ���ѧ������Ҫ��ע�Ը��ơ�����֯�ͽ���������ά�ֺͲ����ṹ�Ļ���ԭ�����漰��������������ۡ�ͼ�������̬����ѧ��������ģ�ͺ��Ŵ��㷨��������1948��롤ŵ�����������Ը����Զ������ۡ����춨���˹�����ѧ�ƻ�����
��һ���˳�����ѧ���ǽ�������Ϊһ������ʽ����Ҫǿ����Ϣ�����ij���ģ�͡���ʵ���ϣ�Ų����ѧ�����˹������������Nils Barricelli����1953-1962�������ڵļ����IAS�ϣ���ʹ����һάԪ���Զ�����ģ����һ���������������������硣�����������������ֹ�����������ʽ�����ݴ���Ϊ�����ھ�������ģ�͵Ĵ�������۲����Խ��������ݻ���ȫò�������ͺ����������ݻ���ռ����Ҫ��λ[1]������˳���ģ�ͺ��㷨���������������Ļ��������������������������˼��Ϊ�˹������춨�����ۺ�ʵ��������
�ڶ�����20����70-90�����������ģ����
��һʱ�ڣ���ѧ���ǿ�ʼ�������ü���ģ�⼼��������Ԫ���Զ����������硢���ֽ���ϵͳ�ȣ����о���Ӧ�ԡ�ӿ�ֵ�����ϵͳ���ԡ�
1987��9�£�����˹�и������٣�Christopher Langton����֯�˵�һ���˹��������飬���־���˹�����ѧ�Ƶĵ����������У�ALife������Ϊ�ԡ����ܴ��ڵ���������life as it could be��[2]���о���������ڵ�����֪������ʽ�Ĵ�ͳ����ѧ�γɶԱȡ����з롤ŵ������Ԫ���Զ��������Լ����������������Ϸ����˹�ٷҡ��ָ���ķ���Լ����ٱ��˵ȷ�չ����ΪAlife����Ҫ������ģ�͡�
���ٶ�����һ��Ԫ���Զ������Բ����ˣ���������ʱ��Ԫ���Զ������߾����ȶ�ģʽ������֯������Ϊ�����������Ե�����Զ���������Ϊ���������ܾ���Դ�ڻ����Ե����Ҳ�Ǹ���ϵͳ���۵���Ҫ˼�롣�ڶ����˳����ڷ�չģ�ͻ����Ͽ�ʼ�����������ģ�⼼������ʵ�����ƶ��о���������Ӧ�Ժ�ӿ�ֵȸ���ϵͳ��������־���˹�����ѧ�ƵĽ�������ʵ���Ŀ�ʼ��
ǰ����Alife���ֵĹ������������Լ�����ģ��ģ�⣬������ץס�����ձ鱾���������������������ţ�������һ�ֶ����������ǡ����ʽ����ͬ�����ʿ���ʵ����ͬ�����������ܹ���[3]����ˣ��������˳��µ���Ҫ����������/����/�����˹�������Soft����

Ȼ������Alife�о��ķ�չ�����У����ɺͷ�˼ʼ�հ��������ҡ�ͬ�ڻ�Ծ����������ѧ�����ء���ɭ��Robert Rosen������1991������˴�������������������Life Itself������Ϊ��������һ������Ч����պϡ���closed to efficient causation���ģ�M,R��ϵͳ*[4]����ϵͳ�������仯������"��"�������ͱ��������ϵͳ��"��"����������ϵͳ��ø�Ͱ�������Ч����պϵĽ�ɫ�����Ǵ���л��Ӧ�ķ����ֲ�������������ѭ������ṹʹ��������ij�������Ժ���������������ζ�������岻��ȫ�ǻ���������ȫͨ��ͼ���ģ�⡣
��M,R��ϵͳ������л��Metabolism��������Repair������л��һ����ƣ���Ϊf����ɴӲ���A������B��ת����������һ����ƣ���Ϊ�����Ӵ�л����B�ϳɴ�л����f������
���ͬʱ���������֪��ѧ�Ҹ�����˹�ơ���������Francisco Varela��Ҳ�������ٵĹ۵�[5]������Ϊ��������龳�Ժ���ʷ�Բ��ɻ�ԭ�����������价���ľ�������Զ����DZ�ڵ�������ģʽ����Ҫ����Щ˼�������˵������˹������˳���
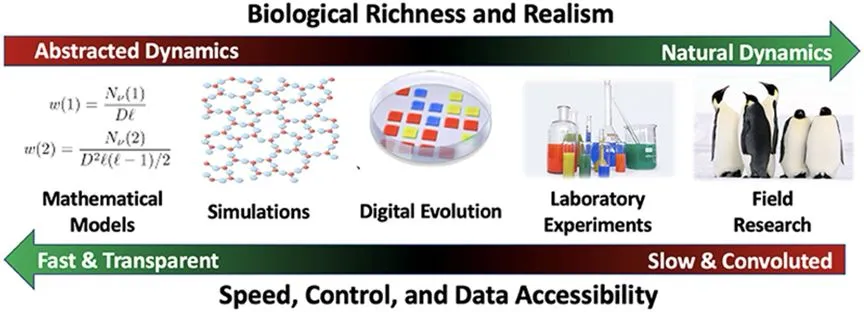
Alife�о��������ף�����ģ�͡�ģ����桢���ֽ���������ʵ�顢ʵ���о��������˴ӳ���ģ�͵���ʵǶ�롢�Լ�����Ӳ��ʪ����Ϸ�ʽ�ķ�չ���̡�[6]
��������20����90�����������Ƕ��ʽ�ݻ���
��һʱ�ڵ��о����ӹ�ע�˹���������ʵ�����Ĺ�ϵ��ǿ����������ʵ�����Ƕ���ԡ������ԡ������Ժ�ӿ���ԡ�1972���1991�꣬�������ֱ�ͺ���������Դ�����Autopoiesis���;������ǣ�Embodied Mind��˼�룬�ֱ�ǿ������ϵͳ�����������������Լ��뻷�����������������Ķ�̬��Ƕ��ʽ��ӿ�ֹ��̡���Щ��Ϊ�˹������о��ṩ���µ������ӽǡ�
���磬�о�����ͼ��ϵͳ�ڲ����롰���ϵ�������������ϵͳ����ƽ��̬�������ͽṹ����������ָ�ݻ��Ͷ���ѧ[7]���Լ��������۲��ߣ���ע�����Ͳ��������뵽ϵͳ���������У�������ʵ���ҽ���ʵ�飬�ƶ���������֯�ݹ�ѭ���в���ͻ�ƺͲ������¹��ܺ���Ϊ����һ����̬ϵͳ����������ķ����ݻ���
���������˹������⣬ʪ����Wet����Ӳ����Hard�����Լ�����˹�������Hybrid������Ϊ��һʱ�ڵĴ��������������-ϸ��������˹���ѧ���ϳ�����ѧ���Ը��ƻ����˵������̽�����о���Χ���˹������չ���˹���̬ϵͳ����ᡣ�������������˹��������о��У���ѧ�ҵ��ص�Ҳ�������־�����ʽ[8]��������̬�ϡ�

EvoSphere�������ݻ�������ϵͳ���ڻ����в���ѡ���ռ��˷����������壬��ͨ�����ߡ����䡱�Ż������ͣ�ͨ��3D��ӡ���������µĻ����˺����[9]
�ܵ����������ǿ���Alife��չ��Ϊ�����ᣬ�����Ǽ���ʵ�ֵķ�ʽ������������ģ�⣨����������Ӳ��ʵ�֣�Ӳ�������ﻯѧʵ�飨ʪ�����������ϵͳ����ϣ����������о����㣬���˹������� �� �˹���̬ �� ����ʽ�ݻ� �� ����������ͼ����
2. �˹���̬�����
����Ȼ���У�����ͨ�����ǹ������ڵģ����磬�����鳤�ද����������������Ⱥ����ʽ����̬ϵͳ�й�������̬ϵͳ����Ϊ�ƶ������ݻ��ĺ���������Ҳ�ɻ��������е��������ﹹ�ɣ�����Ϊһ��������̬�����Ķ�̬ϵͳ����̬ѧ���о����Է�ΪȺ����̬ѧ����̬ϵͳ��̬ѧ�������߷ֱ��ע������Ⱥ��Ļ�������������������֮��Ľ������뵥�������˹������岻ͬ����һ��֧��Alife�о���������õ��˹�ʵ��ļ��ϡ�
����Ȼ��̬Ⱥ���У���ͬ���ֵ���������á���Alife�У��о�������������Ϊij�ֻ����͡������ͻ���̬�ͣ�genotypes��phenotypes��ecotypes��������Ⱥ�䶯̬������������˹��William Reiners��[10]�����ͳһ��ϵͳ��̬ѧ�о���Ҫ���������������������ۿ�ܣ�����ѧ������ѧ���Լ���Ⱥ����û���̬ϵͳ����ͨ�ԡ���connectedness�����ݻ�����Щ��ܿ��Էֱ�����Ϊ���������������������Ϣ��
��������-���ʵ�������ת��������Ӫ������ʳ��������[11]�������ʣ���������ѧѭ������̼ѭ�����ȣ���Ⱥ��������̬ϵͳ����ͨ���������Ⱥ֮�ڵļ�����Ϊ[12]��Ⱥ��֮�������ã��粶ʳ�������������Լ�����̬λ���о���̽����Щ��̬λ���ӿ�֣���ʧ��Ȼ�������[13]��
��ALife�˹���̬�ݻ����о������ۼ�����Ҫ�������Ӷ���ϵͳ��Ԫ���Զ�������������Ϳ����۵ȣ����ֽ���ģ��ƽ̨���ṩ�˼�鸴�ӽ�������̬ϵͳ��̬�Ĺ��ߡ�
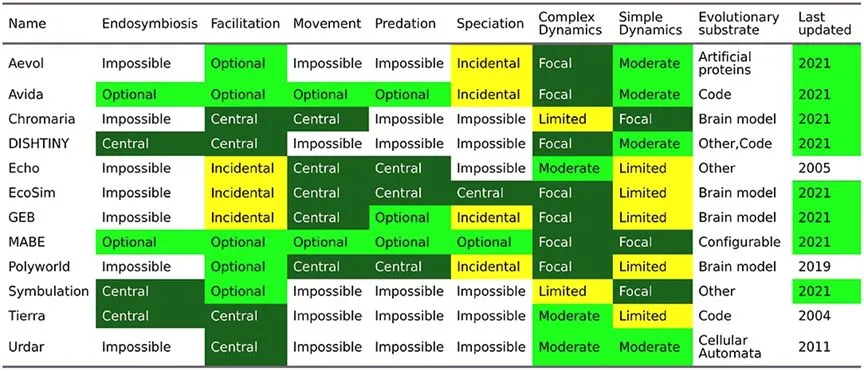
����Alife���ֽ���ģ��ƽ̨��[6]
����һЩģ��ƽ̨����������-������ģ�⡣���磬��Tierra�У�����������Ϊִ��ָ�������CPU���ڣ����������ʱ�䣩������������CPU����ִ��ָ��ı���ֲ���������ˣ���С���������ģ�CPUʹ�ã�������߸���Ч�ʣ���Ϊ������ʩ����ѡ��ѹ������ʹ�䳯�����Ч���ɿ����ݻ�����չ������Avidaƽ̨�У���������ͨ����л��ͬ��������Դ���������������
ALife ��̬�о�ϵͳ����Ȼϵͳ��֮ͬ�����ڣ��о��߿�������Զ����Ȼ�糣��ֵ�IJ���[15]���������趨��ʹ���о����ܹ�̽����Щ����������������ģ���Щ�������¼�����ֻ�ǵ�����������ʽ��żȻ���������ALife�����ܡ��ط�ǰ���ĴŴ�������������ƺͲ���������Ѵ��������зdz���ͬ�ĴŴ�������ǣ����Ӷ������������ȡ�����ı������ԡ�
�˹���̬����Ҳ��Ϊ����Ӳ��ʪ������˹���̬������ʽ��Alife����ƽ̨����Ȼģ�����˹���̬�����л������˹���������ϵͳ����Ϸ�������ڽ��������������̬�����ȡ�Ӳ�˹�������̬��Ҫ�漰������Ⱥϵͳ�Ļ�����ʪ�˹�������̬���Թ���ͨ�����Ṳ���������ɵ���̬Ⱥ�䣬��ִ���������ֽ������̼�ȣ�����̬Ⱥ�䣬���������Ϊ�ϳ���̬ѧ�����ڻ���˹���̬������������ Flora Robotica ��Ŀ[16]������Ŀ������һ��ֲ��ͻ����˹�������̬ϵͳ�������˻�ͨ���ڲ�ͬλ�÷��䲻ͬ��ɫ�Ĺ�������ֲ��������

������ʪ����Ӳ���˹���̬ϵͳ��
�˹���̬ϵͳ������о���ʵ���dz����������磬���Ÿ����Ե����ӣ���̬ϵͳ���Դٽ������Ļ�����[17] �������漰���Ӷᡢ�������Ļ��Ļ����������˶Ը�֪�����������Ե��о���[18]�����⣬���˹��������������塢�˹��������ܵ��о���Ҳ���Թ��Ϊ�˹���̬ϵͳ�������о�[19]��
3. ����ʽ�ݻ�
�������ݻ��Ĺ�ԣ�����ѧ�е�һ�ж�û�����塣��
����������������ѧ�ҵҰ¶������಼��˹����Theodosius Dobzhansky��
�ڲ�����ѡ��������£����з�ֳ���Ŵ��ͱ����������Ⱥ����̬ϵͳ���ᾭ�������ʽ���ݻ��������㷨[20]ʹ�����Ƶĸ����������ⲿ������Ӧ�Ⱥ�����fitness��������ֵ������������������벻���Ľ��[21]��
Ȼ������Ȼ�ݻ��ڶ��������Ҫ���˹�ģ�⸴�ӵö࣬���������鸴���ԡ���Ⱥ��ģ��������������ˮƽ����ת�Ƶ���ѡ��ķ�ֳ���ԡ�������ء������ͱ����Ŵ�ѧ�����á������ͬ�ݻ���Ⱥ֮��Ļ����Լ����ڶ������Ӧ�ȵȷ��档��Щ����Ȼ���ݻ�����������ALife�ݻ��о��ĺ�������[14][22]��
�������ԣ�����֮��������[23]������ʽ�ݻ���Open-Ended Evolution, OEE��[24]����ALife��һ����Ҫ�о�����������������������ϵͳ�������ԡ������뻷�����ù�ϵ�ĸ���������ͬʱ�ṩ��һ�������̽����ģ��DZ��������ʽ�Ŀ����Կռ䡣���о������У������������������Ż���ѧϰ�ͽ������㷨����ˣ�ʵ�ֿ���ʽ�ݻ�Ҳ����˵���˹��������ռ�Ŀ�ꡣ
������һ���ݻ���һ������ʽ�ݻ�ϵͳ��Զ�������뵥һ���ȶ�ƽ�⣬����ӱ�Ի��������[25]�������Ի��������ӡ����������ʽ�ݻ���һ�������˶���߶��ݻ��Ĺ���[26]������ʽ�ݻ������漰������Դ����[27]�������Ժ���֯��γ��ֵ��ش��ݻ�ת��[28]�Լ�Ԫ�ݻ������ݻ����������ݻ����ݻ�[29]�����⡣
Ҫ��������ʽ�ݻ����м�����������[30]��
��1��DZ�ڻ����͵������Ŵ��ռ䡣�Ⲣ����ζ������Ļ����鳤�ȱ���û�����Ƶر仯���������������ͻ����Ŵ��ٶȺ�����Ҳ���ܴ��ڴ����DZ��뵰�ĵ��ػ���ԭ����ν������������Щ����ͨ������ǰ�߿���Ѹ�ٲ��������ԡ�����ζ��ֻ��Ҫͨ���ı���������������Ȩ�صĵ��ؿ��أ����ܺܿ�����γ��µ����֡���һ��������������ձ鹲ͬ���ȣ�LUCA�������ﹲͬ�����Լ�������������
��2��DZ�ڱ���֮����ڶ���ͻ��;��������ζ��DZ������������Ӧ���ܹ������ͬ��ͻ��;�������������˺�������۾��书�����ƣ���ȴ�Ƕ��������ģ���ͺ��������������ͬ�ݻ��İ�������������ѧ��˹ͼ���ء���������Stuart Kauffman��ͨ�������������Դ˽����˿̻��������ֻ�����������������Ϊ���ɽڵ������Լ����ڵ�����K�����ġ���K=2ʱ������ͻ����Ȳ��Dz����㡢�� ��Ҳ������ȫ����Ļ����Ե��״̬����ʱ����ÿһ�������ӾͶ�Ӧһ��ϸ���Ļ���������ͬ�Ļ����飬���Բ�ȡ�˲�ͬ�Ľ���·�����ﵽ�뻷�������ͬһĿ����͡�
��3�������ݻ��Ķ�̬����Ӧ���ۣ�Dynamic adaptive landscape��������ζ����������Χ�Ļ����Ƕ�̬�仯�ġ�һ���棬����������Ⱥ�ݱ䣬����Χ�Ļ���Ҳ������Ϊ���ı䣬һ���涯̬�Ļ�����϶�������Ⱥ����ѡ��ͨ�������Ŵ�������Ӱ������ͺͱ��͵�ʵ�֡�
����ʽ�ݻ�������Ԫ���Զ��������ֽ���ģ��ƽ̨��Tierra��Polyworld��Avida�ȣ��˹���ѧ[31]ģ��ƽ̨Stringmol��[32]�����о���2019��һƪ�о�[33]�������ݻ��ı�����Ϊ�������ռ�Ĺ�ϵ��ȷ�����������͵���ӱ���Լ���Ӧ�Ŀ����ԣ�
̽�������ԣ�Exploratory������ʹ�õ�ǰģ����������ӱ�ԣ�ͨ���漰���������������ϻ����в���ֵ���ġ�������������µĵ�λ�������Ͽ��ܾ������¼�����ļ���Ŀ��
��չ�����ԣ�Expansive����һ����Ҫ�ı�ģ�ͣ�����ʹ�õ�ǰԪģ���д��ڵĸ������ӱ�ԣ��漰��״̬�ռ��з��ִ��������״̬���������漰��֯������ǰδ�����õĻ�ѧ���������ɣ���һ���µı߽�������װ�õij��֡�������г��ij��֣��Լ��Ӿ��й�ϵͳ�ij��֡�
��↑���ԣ�Transformational����һ�������¸����Ҫ�ı�Ԫģ�͵���ӱ�ԡ��ⲻ���漰�����µ��������ɣ�����������֯�㼶�ϵı�Ǩ�����磬�ϳ�һ����ǰδ�ڴ�л��Ӧ��ʹ�õ��»�ѧ���ʡ�������Դ�����ϸ���Լ���ϸ��������Ժ�������ʶ�����ֺͼ��������ڴ��ࡣ��������Ӧ�ڸ���ϵͳ�е�ǿӿ�֡�
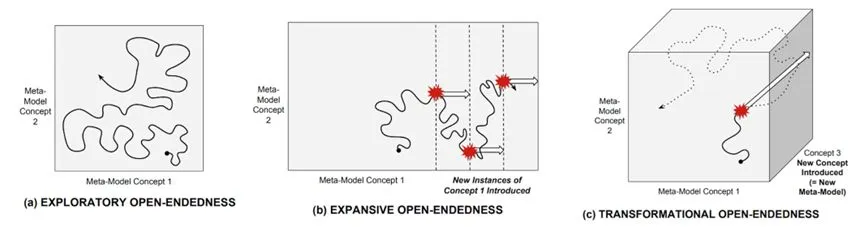

���ֿ����ԣ������ȸ����̬λ����Ӧ�������֣����ϸ����
ͬ�����Կ���ʽ�ݻ��м��ֲ�ͬ�Ķ���ָ��Ҳ���ڼ��[34]�������俪����DZ�������������̬������MODES��ָ��[35]����NKģ�ͺ�Avida���ֽ���ƽ̨�Ȳ�ͬ��ʵ�黷���У���Щָ�����Ч�Ժ��������Ѿ��õ���֤��
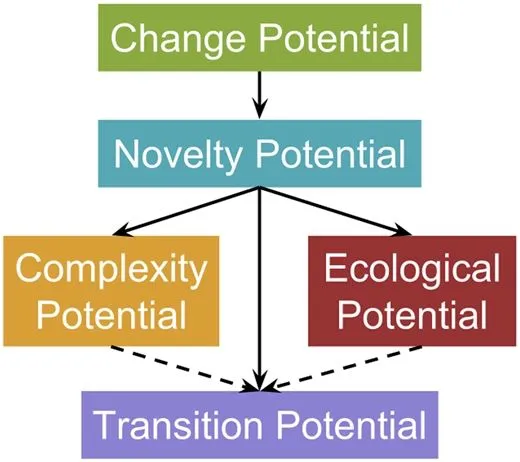
MODES��ָ��֮��Ĺ�ϵ��
�� MODES �У�������DZ����Ҫ�����ܹ����ϵؽ�����Ļ�����Ϣ���ϵ������������У��Ӷ����������ӵ���Ϊ����̬��DZ������������Ⱥ��Ϊһ�������ܹ����պͷ�ӳ������Ϣ����������������������ﻷ������ô����µ���̬λ��Ӫ����ε�������
�ɼ���Ҫʵ�ֳ����Ŀ����Ա����뽫��������̬��������������ݻ������С��ݴˣ�����ʽ�ݻ�ϵͳ������ΪRosen�ģ�M,R��ϵͳ�����⣬�ݴ�����ģ��ChatGPT����������������̽�������ԡ�����ʽ�ݻ����о��Ʊػ�Լ��㴴������AGI�о��ṩ��Ҫ�ķ����ͷ�ʽ��
4. ����
���ݻ��е�һ�У�ֻ���ڼ����Ĺ�â�²������塱����ƪ2021��һƪ���ĵı���[36]��ģ���˶಼��˹�����ݻ�����������������������š�����о�����������-���������������ݻ�[37]��������������ӱ�ԡ�������̬�����Ժ��ƶ����������ش�ת�����Ҫ���ơ���ǰ���������˹�������������˹�����ģ�����ھͶ�Ϥ����㡣��ˡ�������Nick Lane���ڡ�������������Դ����Ҳ��Ϊ���ڹ��������������������Ч�ʣ��ٽ�������������Դ����ʹ�������ݻ��ɵ�����������
������Symbiosis�����Կ��������������Ⱥ֮�䣬�ڸ���������̬�߶��¸�Ϊ���е�Эͬ�ݻ�[38]�Ľ�������ݿռ�λ�ã��������Է�Ϊ�ڹ�����ectosymbiosis���������endosymbiosis�����������ּ�������ϵ����ͨ����������������ƫ�����������Ի�����ƫ�������������Լ�����[39]/��ʳ�����壩[40]����̬ϵͳ����Դ����-��������ѭ���ᵼ����Ⱥ���������ݻ��������γɿ�ι����crossfeeding������һ�����ֵĴ�л���Ϊ��һ�����ֵ���Դ[41]��
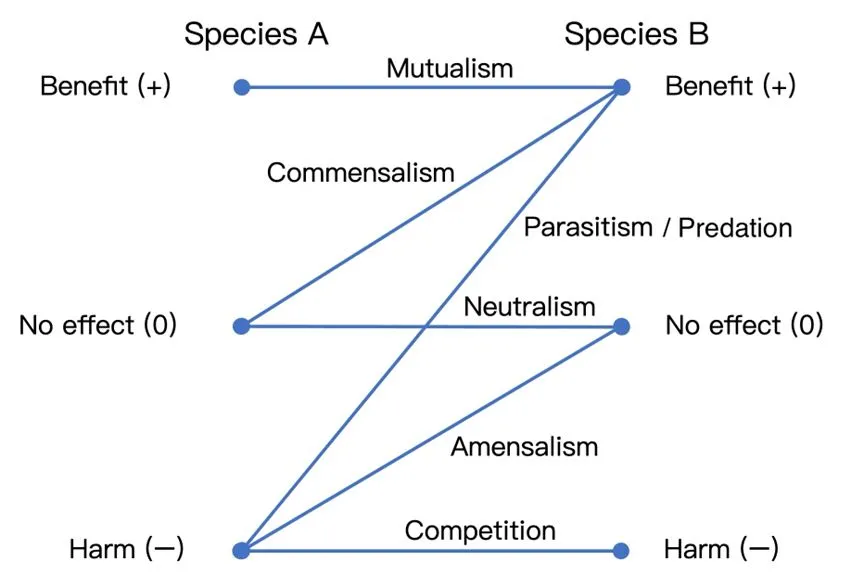
������ϵ[42]�����߽�� https://en.wikipedia.org/wiki/Symbiosis�Լ�[40] Figure 1.1ͼ���ϲ����ơ�
����ͬ��������������Ϸ�����ֽ���ģ��ƽ̨�����о������磬���о����ڿ�����������Ϸ�����Ϸ�չ��Model-Sģ��[43]����ģ�ͳɹ�ģ�����������֯���Դ�������ϸ�����ݻ���������ֳ�ĵ����Լ����ֹ������ںϲ��ԡ��о����֣���ʹ����������Ҳ�����������Ⱥ����Ӧ�ȣ�������֧�ֿ���ʽ�ݻ�����֧�ֹ��������ֽ���ϵͳ��[44]����������ͨ�����ġ�CPUʱ�䡱��Դ������ͷ�չ���ڹ����������������ڲ���Դ������ʵ������ϸ�������������ơ�ͬ�������ֺ����彫���ֻ����飨�����ƴ��룩ת������л��Ϊij���������Ϊ������������ϸ���к������ȡ�Ŵ����벢�ϳɵ����ʵĹ��̡�
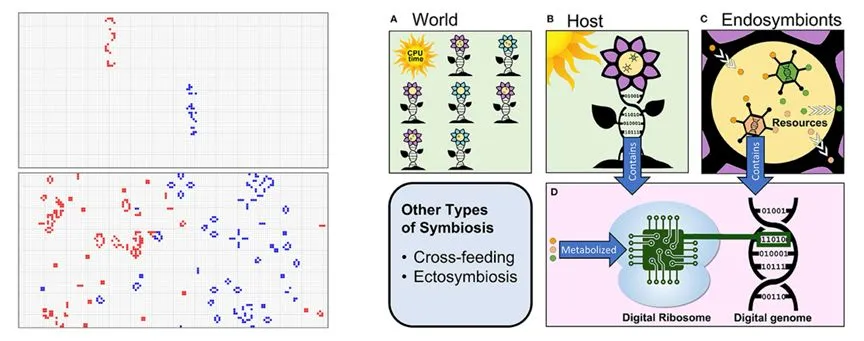
Model-Sģ���������Ϸ�����������㡱[43]�����ֽ���ϵͳ��ģ����ڹ���[44]��
����˵����������Դ�����һ�ֹ��������Һ���˲�ͬ����ĸ����ӡ��ӻ����Ļ���ģ�����Լ�����Ϊ��չ���ͣ�����δ�������������ֵ��ںϿ��ܽ���һ���˻���������ʱ��[45]��
��������ݻ��Ļ��ʻ�����Ȼ������������ô������-��������ݻ����ʺ���̬��������ʲô����Ȼ���������һ������ͻ����������ܽ��л�������ϵͳ�����о�ָ��[46]�������������ķֲ�ʽϵͳ���жԻ�����Ӧ�������ͱ仯���Ŵ����ƺʹﵽ��̬����������������������������������AI������ϣ�����ϵͳ����Ϊ�ȴ�ͳ�����������Ƶ�����֯ϵͳ��������Ϊ�����ڹ����ķֲ�ʽ�������dVM���ṩ��һ���������������Ŀ��Ż�����Ϊͨ���˹����ܣ�AGI���ķ�չ�ṩ֧�֡�
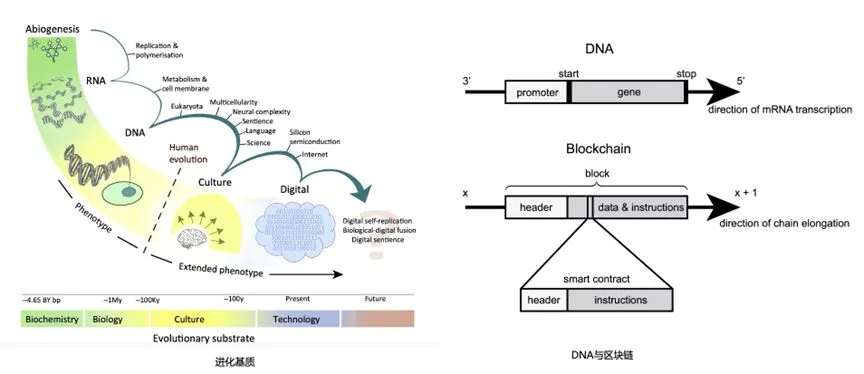
�������ʵ��ݻ�[45]��DNA��������[46]��
�������������������������һ����Ϊ�����������ۣ�Digital Matter Theory���Ĺ۵㣬��������Ϊ��������Ϣ���Ա�����һ���������ʣ���ʵ�����ʣ�����ľͷ����������ơ�ͨ���ھ����������й��е�ģʽ�����Դ���һ�����͵�����������̬��������Ԫ�ر���������Ϊ�����������ʻ��ʺ��ʲ���һ�ֱ��֡���������߶����ӣ������������Ҳ����������һ���ڲ����������䷢չͬ������Ԥ�ڡ���������ϵͳ�У����ǿ��Է��з�����ͬ��Դ/���ң�Non-Arbitrary����ζ����������Ϊ�趨��������������Ϊ����������Ԫ�����ṩ�˵ײ����ͻ�����
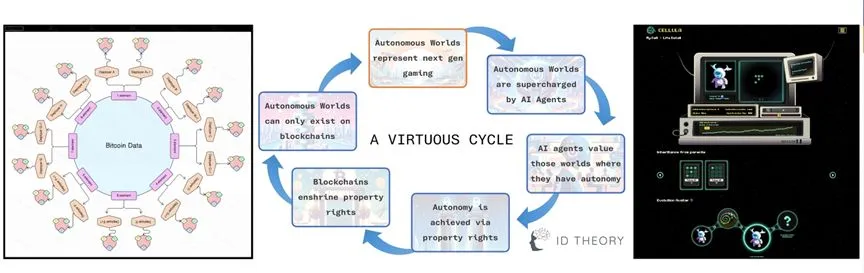
�����������ۡ��������������������ϷCellula[55]��
���⣬ĿǰҲ����һЩ������������Alife��Ŀ���������ϵ��˹�����ģ����Ϸ��Cellula����������������ɵش������ֻ������У����������Լ������ϡ����������������Ϸ�У�����߶Ȱ����ˡ�ʱ�䡱�Ľ�ɫ��ÿ��������������������ʱ����̬ϵͳ�гɳ����������������������ϵ�Alife�������漰������Һ��������������壬����ע����������Ϣ��˫�ݣ������һ�ֵ��͵Ļ��ݹ�����̬�����⣬����һЩ��Ŀ̽����ͨ������������������������ʹ������������ֲ�������ֻ���������Ŀ����ԣ�����terra0�� Flora Robotica��Ŀ��

��ý������ֹ��������ӡ�
��Ȼ���ȽϹ㷺���۵��˻��������˺��˹����ܷ������������Ҫ���������Э�����о����Լ��˻���ֵ����[47]������AI��������[48]������о�����Щ�о��ںܴ�̶���������������������Ĺ۵��Լ�����������Ϊ���Ĺ�����ƿ�����
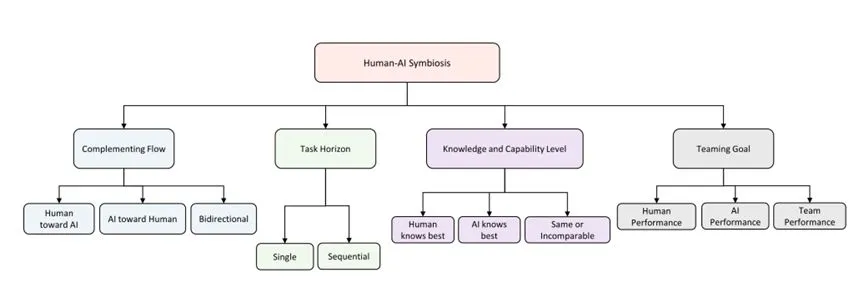
�˻������ķ���ά�ȡ�[49]
5. ��������AI���˹�������δ��
�ع���ʷ���Կ�����AI�����������ڷ����˹���������������������������GOFAIstic��[50]����Alife�����������������౻��Ϊ��������������ΪAI�ķ�չ�ṩ����Ҫ��кͷ�ʽ��
���٣�Geoffrey Hinton�����������ѧϰ���磬ʵ������һ��ģ��������AIϵͳ������Transformer�����Ĵ�ģ��ͨ����ע����������ij�̶ֳ���ģ����������Եļ����ѧϰ���ƣ������Եĺ��������ģ����һ������ѭ������������Transformer����TEM is causal transformer with RNN position encodings����[51]�������� Chatgpt ��OpenAI��ʱ������Ա������ɬ�Ľ�ѵ����The bitter lesson�������Ź�ģ����Scaling Law������ӿ�����ԣ�Emerging properties�� ����ΪֻҪ���������������ݺ������Ĵ�ģ�Ͳ���ʵ��AGI��ǿ��ѧϰ�������෴��ǿ��ѧϰ��RLFH����Դ�ԶԶ�����Ϊ����ѧ�Ľ����������һ�ֻ��ڷ����Ļ�����Ӧ���ƣ�����ʽ�Կ����磨GAN����Դ�Զ��ᄎ���빲������̬ѧ���ơ�
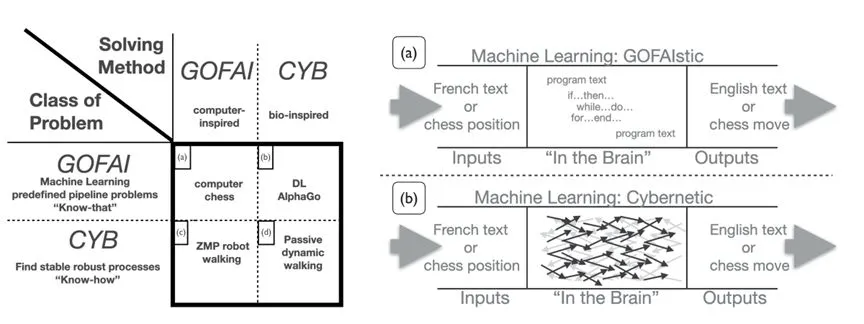
GOFAI���������������������ۣ�AI[52]
���ż���������Alife�о��߲���̽���ӳ���ģ�͵�����ģ�⣬�ٵ�����Ƕ�롢�˹���̬������ʽ�ݻ��ȶ��ַ�ʽ����Щ�����ƶ�AI�Ľ�һ����չ�����磬����������˹�٣�Karl Friston����������ԭ����������������Ƕ��Ԥ��ܹ���I-JEPA��[53]��������AIϵͳ�����ϸ�������ѧԭ��������ǿ���뻷���Ľ��������������ԡ����������Ƿ�ŵ�����ܹ������ٺ�������Ŀ�����㣨Mortal Computation��������Ϊ���۵��˻���ֵ���룬�Լ�����̬�������˹�����[56]���о��������Կ���Alife�ķ�չ��·��Ӱ�ӡ�
��Ȼ��Ŀǰ������ʽAI���ر��Ǵ�����ģ�ͣ�Ҳ�ڶ�Alife�о�������ԶӰ��[54]���������ô�����ģ��̽������IJ�����ӿ�ֺ����ӿ�֡�����ʽ�ݻ����������˻�Э�����˹����������Լ�������ģ�������Ķ�AI������ϵͳ��������˹̹��AIС�����⣬ĿǰҲ���㷨����Ľ�ϣ���������������Ŵ��㷨��������Neuroevolution���������о���ʹ�ü���������[57]��ͨ���Ӿ���������֪����������ͬ��Ͳ�ʳ���������ֲ��ָ���ӿ�ֳ�ר��Ӧ�Բ�ʳ�ߵ�"�־�����"ģ�飬��������������Ե���������ʶ�����ƿ������˹�������ӿ�֡�
������������Alife�ķ�ʽȥ˼����������ǰչ���������У�������������ϵͳ���������������ԭ���������ʵ�־��������Եģ�M,R��ϵͳ��������ģ�ͺ�AI����������ݻ�����↑���ԣ�AI�����ཫ�����γ����ֹ�����ϵ���������ڹ������ǿ�����ι����������ʵ����չ��ʵ�ֽ����Ӱ��δ����������̬��
����δ����Σ�Alife��AI����������Ԫ���棨AR/VR/XR�������֯��չ������һ����Ȼ�������������塢���ֺ����������ڵĹ������硣�������ĺ�����ʱ����������������ʽ������ʵ�嶼���ڹ�������������м����俪���ݻ����ó̡�
�����
[1] D. B. Fogel, "Nils Barricelli - artificial life, coevolution, self-adaptation," in IEEE Computational Intelligence Magazine, vol. 1, no. 1, pp. 41-45, Feb. 2006, doi��10.1109/MCI.2006.1597062.
[2] Alan Dorin, Susan Stepney; What Is Artificial Life Today, and Where Should It Go?. Artif Life 2024; 30��1����1�C15. doi��https://doi.org/10.1162/artl_e_00435
[3] https://medium.com/paul-austin-murphys-essays-on-philosophy/christopher-langton-artificial-life-and-ultra-functionalism-3eb5fa8a92f0
[4]��M,R������л��Metabolism��������Repair������л��һ����ƣ���Ϊf����ɴӲ���A������B��ת����������һ����ƣ���Ϊ�����Ӵ�л����B�ϳɴ�л����f������
[5] Maturana, H. R., & Varela, F. J.��1980��. Autopoiesis and Cognition��The Realization of the Living. D. Reidel Publishing Company.https://doi.org/10.1007/978-94-009-8947-4
[6] Dolson E and Ofria C��2021��Digital Evolution for Ecology Research��A Review. Front. Ecol. Evol. 9:750779. doi��10.3389/fevo.2021.750779
[7] Pattee, H.H.��2012��. Evolving Self-reference��Matter, Symbols, and Semantic Closure. In��LAWS, LANGUAGE and LIFE. Biosemiotics, vol 7. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-5161-3_14
[8] Stepney, S.��2007��. Embodiment. In D. Flower & J. Timmis,��Eds.��In silico immunology��pp. 265�C288��. Springer. https://doi.org/10.1007/978-0-387-39241-7_13
[9] https://towardsdatascience.com/robot-farms-artificial-life-and-second-order-engineering-an-in-depth-interview-with-professor-a524c92ee455
[10] Reiners, William A. ��Complementary Models for Ecosystems.�� The American Naturalist, vol. 127, no. 1, 1986, pp. 59�C73. JSTOR, http://www.jstor.org/stable/2461647. Accessed 27 May 2024.
[11] Lindgren, K., & Nordahl, M. G.��1994��. Artificial food webs. In C. G. Langton��Ed.��, Artificial Life III��Proceedings of the workshop on Artificial Life, Santa Fe, 1992��pp. 73�C103��. Addison-Wesley.
[12] Theraulaz, G., & Bonabeau, E.��1999��. A brief history of stigmergy. Artificial Life, 5��2��, 97�C116. https://doi.org/10.1162/106454699568700
[13] Luo, T.-T., Heier, L., Khan, Z. A., Hasan, F., Reitan, T., Yasseen, A. S., III, Xie, Z.-X., Zhu, J.-L., & Yedid, G.��2018��. Examining community stability in the face of mass extinction in communities of digital organisms. Artificial Life, 24��4��, 250�C276.
[14] Bull, L.��2021��. On the emergence of intersexual selection��Arbitrary trait preference improves female-male coevolution. Artificial Life, 27��1��, 15�C25. https://doi.org/10.1162/artl_a_00335
[15] Dorin, A., Korb, K. B., & Grimm, V.��2008��. Artificial-life ecosystems��What are they and what could they become? In S. Bullock, J. Noble, R. Watson, & M. A. Bedau,��Eds.��Artificial Life XI��pp. 173�C180��. MIT Press.
[16] H. Hamann et al., "Flora Robotica - Mixed Societies of Symbiotic Robot-Plant Bio-Hybrids," 2015 IEEE Symposium Series on Computational Intelligence, Cape Town, South Africa, 2015, pp. 1102-1109, doi��10.1109/SSCI.2015.158.
[17] Marriott, C., Borg, J. M., Andras, P., & Smaldino, P. E.��2018��. Social learning and cultural evolution in Artificial Life. Artificial Life, 24��1��, 5�C9. https://doi.org/10.1162/ARTL_a_00250
[18] Yamauchi, H., & Hashimoto, T.��2010��. Relaxation of selection, niche construction, and the Baldwin effect in language evolution. Artificial Life, 16��4��, 271�C287. https://doi.org/10.1162/artl_a_00006,
[19] https://arxiv.org/abs/2304.05147
[20] De Jong, K.��2005��. Genetic algorithms��A 30-year perspective. In L. Booker, S. Forrest, M. Mitchell, & R. Riolo,��Eds.��Perspectives on adaptation in natural and artificial systems��pp. 1�C32��. Oxford University Press. https://doi.org/10.1093/oso/9780195162929.003.0006
[21] Lehman, J., Clune, J., Misevic, D., Adami, C., Beaulieu, J., Bentley, P. J., Bernard, S., Belson, G., Bryson, D. M., Cheney, N., Cully, A., Donciuex, S., Dyer, F. C., Ellefsen, K. O., Feldt, R., Fischer, S., Forrest, S., Fr��noy, A., Gagne��, C., �� Yosinksi, J.��2020��. The surprising creativity of digital evolution��A collection of anecdotes from the evolutionary computation and Artificial Life research communities. Artificial Life, 26��2��, 274�C306. https://doi.org/10.1162/artl_a_00319
[22] Liard, V., Parsons, D. P., Rouzaud-Cornabas, J., & Beslon, G.��2020��. The complexity ratchet��Stronger than selection, stronger than evolvability, weaker than robustness. Artificial Life, 26��1��, 38�C57. https://doi.org/10.1162/artl_a_00312
[23] Darwin, C. R.��1872��. On the origin of species��6th ed.��. Dent. https://doi.org/10.5962/bhl.title.28875
[24] https://alife.org/encyclopedia/introduction/open-ended-evolution/
[25] Soros, L., & Stanley, K.��2014��. Identifying necessary conditions for open-ended evolution through the artificial life world of Chromaria. In H. Sayama, J. Rieffel, S. Risi, R. Doursat, & H. Lipson��Eds.��, ALIFE 14��Proceedings of the Fourteenth International Conference on the Synthesis and Simulation of Living Systems��pp. 793�C800��. MIT Press. https://doi.org/10.7551/978-0-262-32621-6-ch128
[26] Corominas-Murtra, B., Seoane, L. F., & Sol��, R.��2018��. Zipf��s Law, unbounded complexity and open-ended evolution. Journal of The Royal Society Interface, 15��149��, 20180395. https://doi.org/10.1098/rsif.2018.0395
[27] Cardoso, S. S. S., Cartwright, J. H. E., Čejkov��, J., Cronin, L., De Wit, A., Giannerini, S., Horv��th, D., Rodrigues, A., Russell, M. J., Sainz-D��az, C. I., & T��th, Á.��2020��. Chemobrionics��From self-assembled material architectures to the origin of life. Artificial Life, 26��3��, 315�C326. https://doi.org/10.1162/artl_a_00323
[28] Maynard Smith, J., & Szathm��ry, E.��1995��. The major transitions in evolution. Oxford University Press
[29] Payne, J. L., & Wagner, A.��2019��. The causes of evolvability and their evolution. Nature Reviews Genetics, 20��1��, 24�C38. https://doi.org/10.1038/s41576-018-0069-z
[30] Taylor, T.��2012��. Exploring the Concept of Open-Ended Evolution. https://www.tim-taylor.com/papers/taylor2012exploring.pdf
University of New Mexico, Albuquerque, NM 87131, Ackley, D., & Small, T.��2014��. Indefinitely Scalable Computing = Artificial Life Engineering. Artificial Life 14��Proceedings of the Fourteenth International Conference on the Synthesis and Simulation of Living Systems, 606�C613. https://doi.org/10.7551/978-0-262-32621-6-ch098
[31] Banzhaf, W., & Yamamoto, L.��2015��. Artificial chemistries. MIT Press. https://doi.org/10.7551/mitpress/9780262029438.001.0001
[32] Hickinbotham, S., Clark, E., Stepney, S., Clarke, T., Nellis, A., Pay, M., & Young, P.��2011��. Molecular microprograms. In ECAL 2009, Budapest, Hungary��pp. 297�C304��. Springer. https://doi.org/10.1007/978-3-642-21283-3_37
[33] Tim Taylor; Evolutionary Innovations and Where to Find Them��Routes to Open-Ended Evolution in Natural and Artificial Systems. Artif Life 2019; 25��2����207�C224. doi��https://doi.org/10.1162/artl_a_00290
[34] Susan Stepney, Simon Hickinbotham; On the Open-Endedness of Detecting Open-Endedness. Artif Life 2023; doi��https://doi.org/10.1162/artl_a_00399
[35] Emily L. Dolson, Anya E. Vostinar, Michael J. Wiser, Charles Ofria; The MODES Toolbox��Measurements of Open-Ended Dynamics in Evolving Systems. Artif Life 2019; 25��1����50�C73. doi��https://doi.org/10.1162/artl_a_00280
[36] Hickinbotham Simon J., Stepney Susan and Hogeweg Paulien 2021Nothing in evolution makes sense except in the light of parasitism��evolution of complex replication strategiesR. Soc. Open Sci.8210441
[37] Vostinar, A. E., & Ofria, C.��2018��. Spatial structure can decrease symbiotic cooperation. Artificial Life, 24��4��, 229�C249. https://doi.org/10.1162/artl_a_00273
[38] Weisbuch, G., Duchateau, G. Emergence of mutualism��Application of a differential model to endosymbiosis. Bltn Mathcal Biology 55, 1063�C1090��1993��. https://doi.org/10.1007/BF02460700
[39] Hickinbotham, S., Stepney, S., & Hogeweg, P.��2021��. Nothing in evolution makes sense except in the light of parasitism��Evolution of complex replication strategies. Royal Society Open Science, 8��8��, 210441. https://doi.org/10.1098/rsos.210441
[40] Bronstein, Judith L.��ed.��, 'The study of mutualism', in Judith L. Bronstein��ed.��, Mutualism��Oxford, 2015; online edn, Oxford Academic, 17 Sept. 2015��, https://doi.org/10.1093/acprof:oso/9780199675654.003.0001
[41] Crombach A, Hogeweg P. Evolution of resource cycling in ecosystems and individuals. BMC Evol Biol. 2009 Jun 1;9:122. doi��10.1186/1471-2148-9-122. PMID��19486519; PMCID��PMC2698886.
[42] ���߽�� https://en.wikipedia.org/wiki/Symbiosis �Լ�[40] Figure 1.1ͼ���ϲ�����.
[43] Turney PD. Evolution of Autopoiesis and Multicellularity in the Game of Life. Artif Life. 2021 Jun 11;27��1��:26-43. doi��10.1162/artl_a_00334. PMID��34529755.
[44] Vostinar AE, Skocelas KG, Lalejini A and Zaman L��2021��Symbiosis in Digital Evolution��Past, Present, and Future. Front. Ecol. Evol. 9:739047. doi��10.3389/fevo.2021.739047
[45] Gillings MR, Hilbert M, Kemp DJ. Information in the Biosphere��Biological and Digital Worlds. Trends Ecol Evol. 2016 Mar;31��3��:180-189. doi��10.1016/j.tree.2015.12.013. Epub 2016 Jan 8. PMID��26777788.
[46] Abramov, O., Bebell, K.L. & Mojzsis, S.J. Emergent Bioanalogous Properties of Blockchain-based Distributed Systems. Orig Life Evol Biosph 51, 131�C165��2021��. https://doi.org/10.1007/s11084-021-09608-1
[47] https://arxiv.org/abs/2310.19852
[48] https://arxiv.org/abs/2202.09447
[49] https://arxiv.org/pdf/2103.09990
[50] https://zh.wikipedia.org/wiki/GOFAI
[51] https://arxiv.org/abs/2112.04035
[52] Harvey I. Motivations for Artificial Intelligence, for Deep Learning, for ALife��Mortality and Existential Risk. Artif Life. 2024 Feb 1;30��1��:48-64. doi��10.1162/artl_a_00427. PMID��38346273.
[53] https://arxiv.org/abs/2301.08243
[54] https://direct.mit.edu/artl/article/29/2/141/116359/Editorial-What-Have-Large-Language-Models-and
[54]https://doi.org/10.1162/isal_a_00431
[55] https://medium.com/p/68c29c1e374e
https://medium.com/p/2f1853f8a1d7
https://medium.com/p/1d81efda13��
[56] Han BA, Varshney KR, LaDeau S, Subramaniam A, Weathers KC, Zwart J. A synergistic future for AI and ecology. Proc Natl Acad Sci U S A. 2023 Sep 19;120��38��:e2220283120. doi��10.1073/pnas.2220283120. Epub 2023 Sep 11. PMID��37695904; PMCID��PMC10515155.
[57] Kevin Godin-Dubois, Sylvain Cussat-Blanc, Yves Duthen; July 18�C22, 2021. "Spontaneous modular NeuroEvolution arising from a life/dinner paradox." Proceedings of the ALIFE 2021��The 2021 Conference on Artificial Life. ALIFE 2021��The 2021 Conference on Artificial Life. Online.��pp. 95��. ASME. https://doi.org/10.1162/isal_a_00431